Hoofdstuk 1.Inleiding
1.1.Inleiding
Het Sigmaplan werd in 1977 opgezet om het Scheldebekken tegen overstromingen te beschermen. Na de actualisatie in 2005 is daar een tweede doelstelling bijgekomen: de Scheldenatuur laten herademen. Maar welk effect hebben het Sigmaplan, andere ingrepen en evoluties precies op het ecosysteem? Hoe is het met de waterkwaliteit van de Schelde gesteld? Hoe evolueren de troebelheid en de morfologie? Met uitgebreide onderzoeks- en monitoringsprogramma’s zoals OMES (Onderzoeksprogramma Milieu-Effecten Sigmaplan) volgen de Vlaamse waterbeheerders en wetenschappers de waterkwaliteit, de waterkwantiteit en de flora en fauna in het Schelde-estuarium van dichtbij op. Dit rapport stelt u de resultaten van de waterkwaliteit voor en schetst de evolutie die de rivier doormaakt.
1.2.Ontstaan van OMES
OMES werd opgestart om de effecten van het Sigmaplan op het milieu in kaart te brengen. De stormvloed van 3 januari 1976 en de daar op volgende overstromingen gaven aanleiding tot de uitvoering van dit omvangrijke plan dat het gehele Zeescheldebekken dient te beschermen tegen overstromingen. Het Sigmaplan moest de Vlaamse tegenhanger vormen van het Nederlandse Deltaplan, maar kwam slechts langzaam op gang.
Na een nieuwe storm in 1994, met de hoogste waterstanden ooit gemeten op de Zeeschelde, werd een nood- en urgentieprogramma goedgekeurd. De uitvoering hiervan diende te passen binnen een integrale visie op het beheer van de waterlopen. Dit leidde tot een Algemene Milieu-Impact studie voor het Sigmaplan en daarop aansluitend tot een Onderzoek Milieu-Effecten Sigmaplan, OMES genaamd.
Het OMES-project had tot doel de biogeochemische kennis van de Zeeschelde te actualiseren en in belangrijke mate uit te breiden. Een uitgebreid monitoringprogramma werd opgestart: “Onderzoek naar de gevolgen van het Sigmaplan, baggeractiviteiten en havenuitbreiding in de Zeeschelde op het milieu”, kortweg OMES. Een estuarium is echter een zeer complex systeem dat door verschillende (menselijke) factoren wordt beïnvloed. Monitoring moet dan ook gebeuren vanuit verschillende onderzoeksdomeinen. In deze nieuwe visie waarbij veiligheid, toegankelijkheid en natuurlijkheid samen worden aangepakt, is een goede kennis van het estuariene ecosysteem van de Schelde onontbeerlijk. OMES is een belangrijk instrument geworden bij het wetenschappelijk onderzoek in het estuarium. Monitoring wordt echter pas waardevol wanneer de metingen systematisch, over lange periode worden volgehouden. Dan pas kunnen langetermijneffecten en trends onderscheiden worden van kortetermijnvariaties. Belangrijk is dus een dataset te creëren zonder grote hiaten. OMES kan terugblikken op een grote, consistente dataset: sinds december 1995 werd maandelijks op vaste plaatsen, volgens een vast schema bemonsterd. Op enkele uitzonderingen na zijn er nooit onderbrekingen geweest in het staalnamepatroon.
1.3.OMES - MONEOS - Evaluatiemethodiek
In het voorjaar van 2008 werd MONEOS voorgesteld: een programma voor geïntegreerde monitoring van het Schelde‐estuarium in Nederland en Vlaanderen onder coördinatie van de Vlaams-Nederlandse Scheldecommissie (VNSC). Het MONEOS programma beschrijft een monitoring die wenselijk is om evoluties in de Schelde te beschrijven en oorzaak‐gevolg relaties te achterhalen. Dat is essentieel om op een wetenschappelijk verantwoorde manier het estuarium te beheren. Het plan omvat verschillende aspecten van het Schelde ecosysteem, gaande van hydro‐ en morfodynamiek, diversiteit habitats, fysico‐chemie, ecologisch functioneren en diversiteit soorten. OMES vervult in dit kader een belangrijk deel van de fysico‐chemische en ecologsiche monitoring voor de Zeeschelde.
In het kader van MONEOS werden de bestaande monitoringsactiviteiten onder de loep genomen. Hieruit bleek er een opportuniteit te zijn om het OMES programma en de VMM activiteiten beter op elkaar af te stemmen. In 2009 werd het OMES programma afgestemd op de aanbevelingen uit MONEOS. VMM en OMES bemonsteren nu gezamenlijk de waterkwaliteit van de Schelde, waarbij de klemtoon binnen OMES op de ecologie ligt, bij VMM meer op toxicologie (onder andere zware metalen, polluenten). Een overlap in parameters werd gereduceerd en er kwam een nauwe samenwerking en uitwisseling van gegevens. De staalnamecampagnes werden uitgebreid naar de Rupel.
Als vervolg op het MONEOS, werd door Deltares en Universiteit Antwerpen in opdracht van de Vlaams-Nederlandse Scheldecommissie de Evaluatiemethodiek Schelde-estuarium (Maris et al., 2014) ontwikkeld om op basis van die MONEOS dataset het functioneren van het Schelde-estuarium te evalueren. Voordien bestonden er natuurlijk reeds verschillende beoordelingskaders (o.a. inzake veiligheid, Kaderrichtlijn Water, Habitatrichtlijn), maar een overkoepelende evaluatie van het systeem ontbrak. Ter ondersteuning van beheer en beleid is echter een overkoepelende evaluatie gewenst. Daarom werd de Evaluatiemethodiek ontwikkeld met als uitgangspunten de beleidsdoelstellingen uit de LangeTermijnVisie (LTV) en het goed functioneren van het ecosysteem. De basis is dus niet een historische of ongerepte referentie, wel een systeem dat goed functioneert.
De Evaluatiemethodiek is opgebouwd rond de drie hoofdfuncties van de LTV: Veiligheid, Toegankelijkheid en Natuurlijkheid. Vanuit deze functies is een beperkte set communicatie-indicatoren ontwikkeld. Voor de evaluatie van Natuurlijkheid zijn vijf communicatie-indicatoren opgesteld: Plaat- en geulsystemen, Leefomgeving, Waterkwaliteit, Ecologisch functioneren en Flora & Fauna. Het OMES monitoringsprogramma zal in hoofdzaak data aanleveren voor de indicator Waterkwaliteit.
Elke communicatie-indicator is uitgewerkt in een zelfstandig hoofdstuk in de evaluatiemethodiek, waarbij de beoordeling een getrapte aanpak volgt: de piramide-aanpak (Figuur 1-1). Bovenaan de piramide staat de communicatie-indicator. Deze wordt beoordeeld door de onderliggende niveaus te doorlopen.
Niveau 1: Rechtstreeks onder de communicatie-indicator staan de toetsparameters: een beperkte maar toch volledige set parameters waarmee ondubbelzinnig de toestand of trend van de communicatie-indicator geëvalueerd kan worden.
Niveau 2: Elke toetsparameter is opgebouwd uit één of meerdere rekenparameters. Voor elke rekenparameter is een duidelijk toetsingscriterium opgesteld. Alle rekenparameters samen bepalen of de ontwikkelingen van een toetsparameter gunstig of ongunstig zijn voor het functioneren van het systeem.
Niveau 3: Onderaan de piramide staat een set verklarende parameters. Verklarende parameters dragen vooral bij aan het begrijpen van de waargenomen ontwikkelingen en minder aan het beoordelen van de ontwikkeling. De verklarende parameters zelf zijn immers niet onafhankelijk te beoordelen.
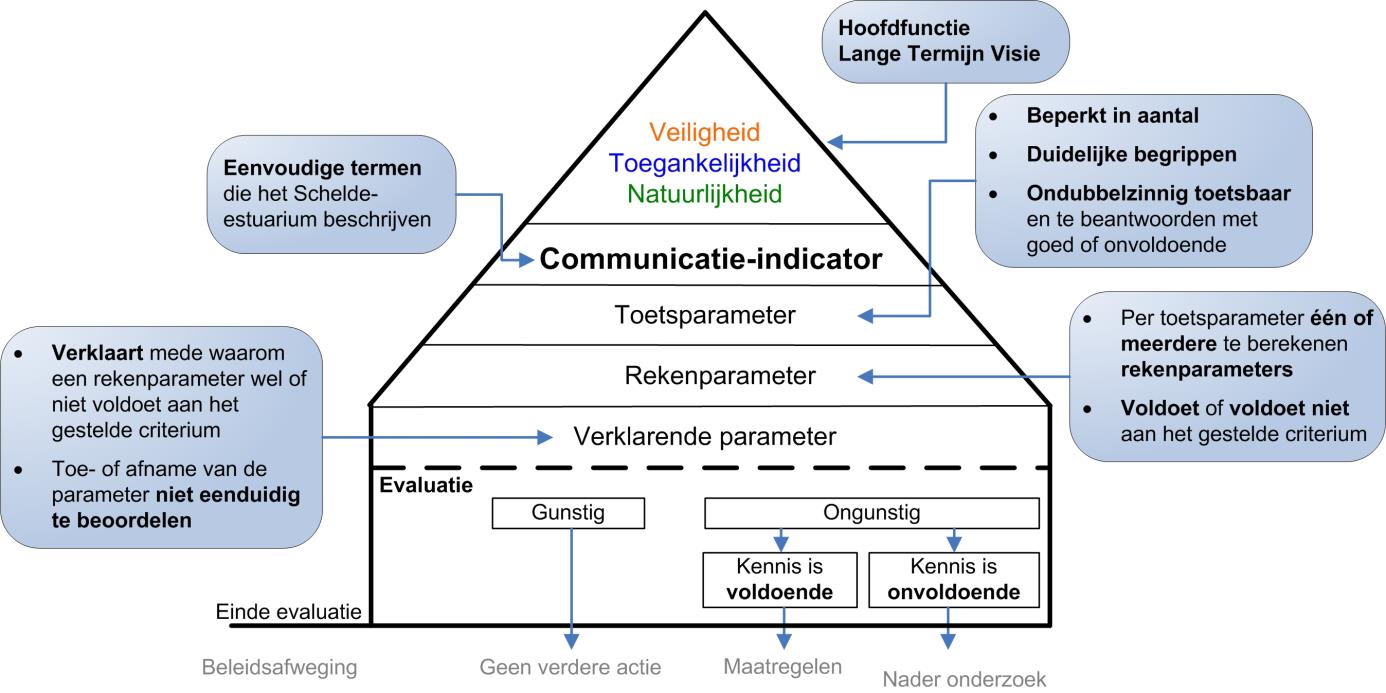
Figuur 1-1 Schematische weergave ‘piramide-aanpak’
OMES maakt deel uit van het grensoverschrijdende monitoringsprogramma MONEOS, de geïntegreerde monitoring van het Schelde-Estuarium. Het monitoringprogramma bouwt verder op bestaande monitoringsactiviteiten en streeft naar integratie en afstemming over de grenzen heen.
Binnen deze OMES rapportage zal waar mogelijk de Evaluatiemethodiek toegepast worden. Een volledige beoordeling van een piramide is echter niet mogelijk, aangezien hiervoor input vereist is vanuit andere disciplines die niet in OMES vervat zitten. Maar verschillende reken-, toets- of verklarende parameters kunnen wel bepaald en beoordeeld worden.
1.4.Huidige OMES programma
Binnen het OMES project worden drie types monitoring uitgevoerd: systeemmonitoring, onderzoeksmonitoring en projectmonitoring.
1.4.1.Systeemmonitoring
De systeemmonitoring is een langlopend programma dat alle basisparameters voor het goed functioneren van het Schelde‐ecosysteem opvolgt. De maandelijkse OMES monitoring op de vaste stations en een 13-uurs opvolging in de Zeeschelde vormen belangrijke onderdelen van deze systeemmonitoring voor waterkwaliteit in het estuarium. Stalen over de ganse lengteas van de Zeeschelde en Rupel worden onderzocht op ruim 50 parameters, gaande van zuurstof en nutriënten over profielen van zwevende stoffen en debieten tot fotosynthese en plankton. De resultaten geven over langere tijd een schat aan informatie over hoe de ecologische toestand van onze getijdenrivieren evolueert.
Elke OMES monitoringscampagne wordt opgedeeld in 3 vaardagen, normaal telkens een maandag (Beneden-Zeeschelde), dinsdag (Rupelbekken en Lippenbroek-Baasrode) en woensdag (Boven-Zeeschelde) (zie Figuur).
Dag 1: vertrek te Antwerpen, afvaart tot station Grens en vanaf daar opvarend meten tot Temse. Terugvaart naar Antwerpen.
Dag 2: vertrek te Boom, opvaren tot Dijle en Netemonding. Afvarend meten tot Rupelmonde. Vanaf daar opvarend meten tot Baasrode en terugkeer per minibus naar Boom.
Dag 3: Vertrek te Dendermonde, opvarend meten tot Melle. Terugvaart naar Dendermonde.
Een overzicht van de stations en de vaardagen wordt gegeven bij Materialen en Methoden.
In het winterhalfjaar (oktober – maart) wordt 1 monitoringscampagne per maand georganiseerd, telkens samen met VMM. In het zomerhalfjaar (april – september) wordt de monitoring uitgebreid tot 2 campagnes per maand, om snel fluctuerende ecologische fenomenen zoals algenbloei adequaat te kunnen registreren. VMM neemt niet deel aan deze extra campagnes: voor een goede opvolging van toxische stoffen volstaat de maandelijkse staalname.
In het kader van MONEOS wordt ook met Nederland een optimale afstemming nagestreefd. In het verleden betekende dit dat de vaartochten van OMES en NIOZ perfect op elkaar afgesteld werden. In 2014 werden echter de maandelijkse campagnes van NIOZ afgevoerd, en wordt de monitoring van de Westerschelde enkel door Rijkswaterstaat verzorgd. Voor staalname en analyse is nog steeds een goede afstemming tussen Vlaanderen en Nederland aanwezig. De frequentie van staalname, zowel in ruimte als tijd, is nu echter lager in Nederland, waardoor de spatio-temporele afstemming minder gunstig geworden is.
1.4.2.Onderzoeksmonitoring
De onderzoeksmonitoring omvat de gedetailleerde opvolging van pilootprojecten om meer wetenschappelijk inzicht te verwerven. In Lippenbroek loopt zo’n onderzoeksmonitoringsproject waarbij kennis vergaard wordt over de ontwikkeling van gecontroleerde overstromingsgebieden met gecontroleerd gereduceerd getij.
1.4.3.Projectmonitoring
De OMES monitoring in Bergenmeersen, Burchtse Weel en Bazel zijn voorbeelden van projectmonitoring. Met een programma voor projectmonitoring wordt gedurende de eerste drie tot zes jaar na de inrichting van deze gebieden nagegaan in hoeverre ze voldoen aan de gestelde verwachtingen en of ze al dan niet in goede richting evolueren. Door de vinger aan de pols te houden, worden mogelijke problemen sneller gedetecteerd. Dat maakt adaptief beheer en bijsturing mogelijk. Het MONEOS‐rapport omschrijft projectmonitoring als die monitoring die − aanvullend op de reguliere systeemmonitoring − wordt uitgevoerd om bepaalde parameters met verhoogde frequentie te meten, beperkt in tijd en ruimte. Na een periode van drie tot zes jaar volgt een evaluatie. Als alles naar wens verloopt, kan de projectmonitoring worden afgebouwd en wordt het gebied opgenomen in de reguliere systeemmonitoring.
1.5.Partners
De huidige OMES monitoring wordt gefinancierd door W&Z (Waterwegen en zeekanaal NV, Vlaamse Overheid; Beleidsdomein Mobiliteit en Openbare Werken). Via een openbare aanbesteding werd de OMES monitoring gegund aan de Universiteit Antwerpen, die een interdisciplinair team van wetenschappers van verschillende universiteiten en instituten samenstelde voor deze opdracht. De taken binnen OMES vereisen immers experts vanuit verschillende wetenschappelijke disciplines:
- UAntwerpen - Universiteit Antwerpen, Onderzoeksgroep ecosysteembeheer (ECOBE)
De UAntwerpen is coördinator van het OMES programma. Voor de systeemmonitoring leidt ze de campagnes en staat in voor de monitoring van de basiswaterkwaliteit. De onderzoeks- en projectmonitoring neemt de UAntwerpen ook voor haar rekening.
- VUB - Vrije Universiteit Brussel - Laboratorium Analytische-, Milieu- , en Geochemie
De VUB staat in voor de opvolging van de koolstofcyclus, zowel bij de systeem- als onderzoekmonitoring.
- IMDC - International Marine and Dredging Consultants
Dit private studiebureau vult het OMES team aan omwille van de sterke expertise inzake het meten van zwevende stof- en stroomsnelheidsprofielen met de ADCP techniek.
- UGent - Universiteit Gent - Protistologie en aquatische ecologie (PAE)
De UGent is de specialist bij uitstek voor de monitoring van het fytoplankton (plantaardige plankton).
- NIOZ - Koninklijk Nederlands Instituut voor Onderzoek der Zee
Dit Nederlandse kennisinstituut voert metingen van primaire productie uit: zij bepalen de productiviteit en groeikarakteristieken van de aanwezige algen in de Zeeschelde.
- ECOLAB - Université Paul Sabatier (Toulouse): Laboratoire d'Ecologie Fonctionnelle
Voor de determinatie en analyse van het aanwezige zoöplankton (dierlijk plankton), wordt beroep gedaan op ECOLAB, een Franse onderzoeksgroep met Vlaamse roots.
- VLIZ - Vlaams Instituut voor de Zee
Het Vlaams Instituut voor de Zee bouwde een sterke reputatie op als datacentrum voor mariene en bij uitbreiding estuariene wetenschappen. Zij zijn bijgevolg de partner bij uitstek om ook voor OMES het databeheer in goede banen te leiden.
1.6.OMES rapport
Dit OMES rapport beschrijft de evolutie van het ecosysteem in de waterkolom (pelagiale ecosysteem) van de Zeeschelde en Rupel. Hiervoor wordt de Evaluatiemethodiek als leidraad gebruikt, en waar mogelijk worden het estuarium geëvalueerd zoals beschreven in de methodiek.
In de Evaluatiemethodiek Schelde-estuarium wordt het pelagiale ecosysteem beoordeeld in de piramide Waterkwaliteit aan de hand van vier toetsparameters: zuurstof, nutriënten, algen en toxische stoffen. Aangezien de meting van toxische stoffen niet tot OMES behoort (VMM neemt deze taak op), zal deze laatste toetsparameter niet behandeld worden. Zuurstof (inclusief biochemische zuurstofvraag en de verschillende koolstofverbindingen) en de nutriënten stikstof, fosfor en silicium worden in dit rapport in hoofdstuk 43 ‘Biologische Waterkwaliteit’ besproken en waar mogelijk ook getoetst aan de criteria uit de Evaluatiemethodiek. Ook pH, verklarende parameter in de Evaluatiemethodiek, wordt hierbij betrokken. Verklarende parameters temperatuur, debiet (verblijftijd) en chloride (zout) komen aan bod in het hoofdstuk 34 ‘Klimatologisch overzicht’. Algen worden besproken in een volgend hoofdstuk 6 ‘Fyto- en zoöplankton’. Ook ‘Lichtklimaat en zwevende stof’ vormen een afzonderlijk hoofdstuk 5.
Staalnamelocaties en een overzicht van de gebruikte methodiek vormen het onderwerp van hoofdstuk 2: Materialen en methoden.
Hoofdstuk 2.Materialen en Methoden
2.1.Staalname
De OMES campagnes hebben een traditie van tijonafhankelijke staalname, waardoor steeds wordt bemonsterd in verschillende fasen van het getij. Echter, sommige stoffen vertonen grote schommelingen met het getij. Zwevende stof bijvoorbeeld wordt bij hoge stroomsnelheden opgewerveld en zal uitzakken rond kentering. Om de staalname te kaderen in de fase van het getij, wordt daarom steeds stroomsnelheid en -richting mee gemeten. Ook opgeloste stoffen kunnen een sterke tijvariatie kennen, wanneer er voor die stof een sterke longitudinale gradiënt bestaat, met andere woorden wanneer die stof een sterke toename of afname kent in op- of afwaartse richting. Om tijafhankelijke fluctuaties beter te interpreteren, worden daarom jaarlijks 3 dertienuursmetingen (= volledige tijcyclus) uitgevoerd op 3 locaties in het estuarium.
2.1.1.Estuariene campagnes
De locaties in het estuarium, de estuariene stations (Tabel 2-1, Figuur 2-1), worden per schip bemonsterd. Zo kan in het midden van de stroom een representatief monster genomen worden. Om alle stations onder min of meer gelijke weersomstandigheden te bemonsteren, worden deze binnen een tijdsspanne van drie dagen bemonsterd. De monstername is opgedeeld in drie opeenvolgende vaardagen, met telkens vertrek tussen 8 en 9 uur. Op deze wijze kunnen alle station bij voldoende daglicht bemonsterd worden, wat naar veiligheid een voordeel is, maar voor de metingen van het lichtklimaat een must.
Tabel 2-2 geeft een overzicht van alle campagnedagen in 2015. Wanneer er in het zomerhalfjaar twee campagnes per maand zijn, is de eerste steeds in samenwerking met VMM; aan de tweede campagne neemt VMM niet deel. In het winterhalfjaar is er 1 maandelijkse campagne, steeds samen met VMM.
Tijdens alle campagnes werd telkens 1 diepteprofiel gemeten van de stroming, zwevende stof en troebelheid. Een overzicht van opmerkingen tijdens het verloop van de meetcampagnes is ook terug te vinden in Tabel 2-2.
De grenzen van het estuariene systeem, de zogenaamde boundaries (Figuur 2-1), worden binnen dezelfde periode van drie dagen bemonsterd, tevens in nauwe samenwerking met VMM. Ook hier worden tijonafhankelijk schepstalen genomen. Deze plaatsen worden echter niet meer per schip bemonsterd. De locaties zijn zo gekozen dat een representatief staal genomen kan worden van de waterloop, meestal vanop een kade, ponton of brug.
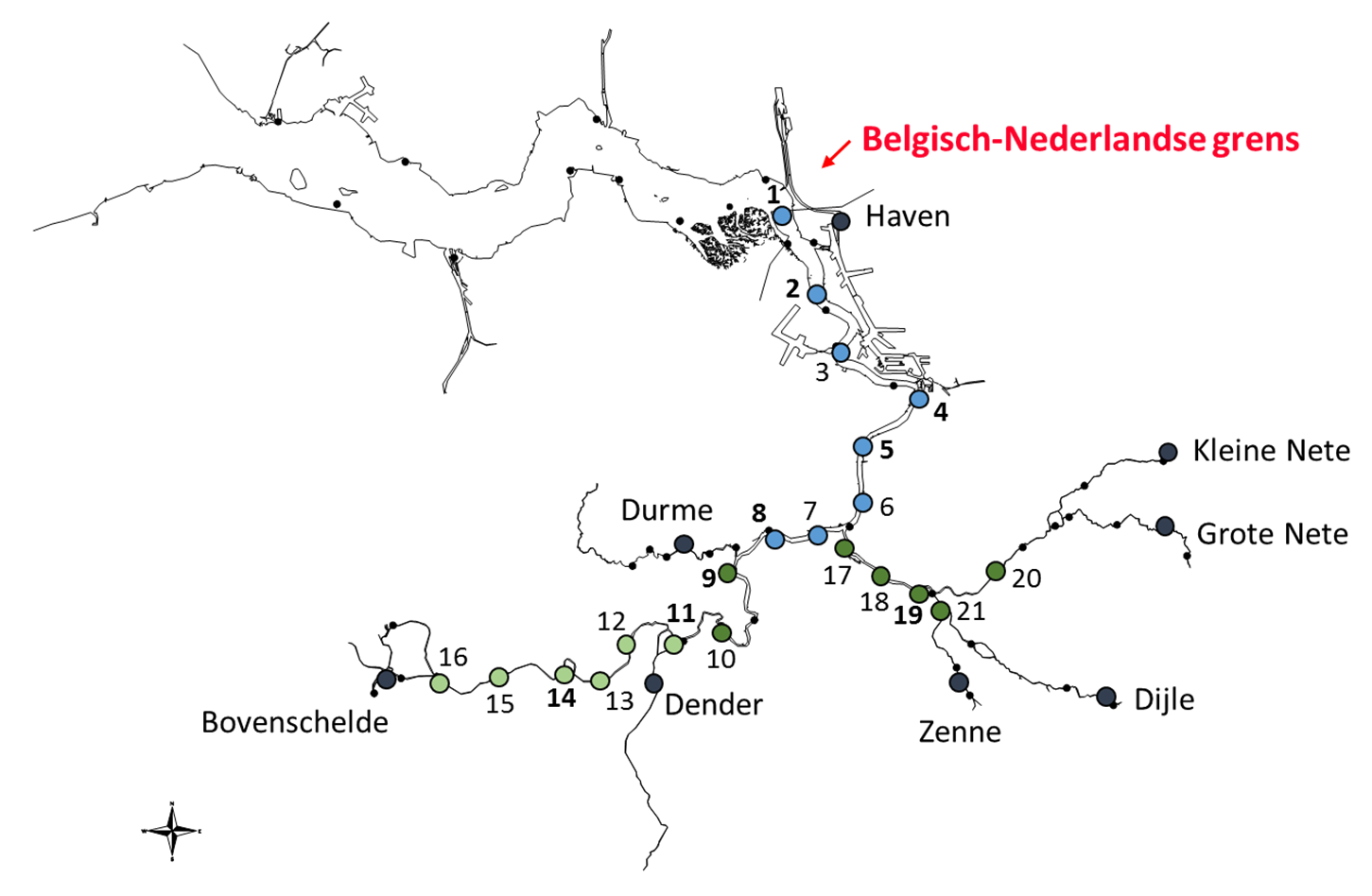
Figuur 2-1 Overzicht van de locaties voor de systeemmonitoring in OMES. Donkergrijze stippen duiden de boundaries aan, met vermelding van de naam. De gekleurde stippen duiden de estuariene stations aan (blauw: monstername op vaardag 1, dondergroen op vaardag 2, lichtgroen op vaardag 3; zie Tabel 2-1 voor nummering). Op stations met nummering in vet worden dwarsprofiel opgenomen.
Tabel 2-1 monitoringsstations in het OMES programma met aanduiding van de staalnamedag en de afstand tot de monding (Vlissingen, volgens OMES databank)
| station | Herkenningspunt | rivier | km t.o.v. monding | staalname | |
| 1 | Grens | boei 87 | Zeeschelde | 58 | dag 1 |
| 2 | Liefkenshoek | boei 92, Liefkenshoektunnel | Zeeschelde | 63,5 | dag 1 |
| 3 | Melsele | Punt van Melsele | Zeeschelde | 72 | dag 1 |
| 4 | Antwerpen | ter hoogte van het Steen | Zeeschelde | 79 | dag 1 |
| 5 | Kruibeke | Veerponton | Zeeschelde | 85 | dag 1 |
| 6 | Bazel | Veerponton | Zeeschelde | 89 | dag 1 |
| 7 | Steendorp | ter hoogte van de kerk | Zeeschelde | 94 | dag 1 |
| 8 | Temse | Afwaarts spoorbrug | Zeeschelde | 98,5 | dag 1 |
| 9 | Lippenbroek | ter hoogte van GOG-GGG | Zeeschelde | 103,5 | dag 2 |
| 10 | Baasrode | Veerponton | Zeeschelde | 113 | dag 2 |
| 11 | Dendermonde | afwaarts brug | Zeeschelde | 121 | dag 3 |
| 12 | Sint Onolfs | ‘bocht van Damme’ | Zeeschelde | 125 | dag 3 |
| 13 | Appels | veerponton | Zeeschelde | 128 | dag 3 |
| 14 | Uitbergen | Afwaarts baanbrug | Zeeschelde | 138 | dag 3 |
| 15 | Wetteren | Afwaarts baanbrug | Zeeschelde | 145 | dag 3 |
| 16 | Melle | Afwaarts baanbrug | Zeeschelde | 151 | dag 3 |
| 17 | Niel | Loskade | Rupel | 94 | dag 2 |
| 18 | Boom | veer | Rupel | 98 | dag 2 |
| 19 | Terhagen | kerk | Rupel | 102 | dag 2 |
| 20 | Beneden-Nete | Tussen Brug E19 – N1 | Nete | 105 | dag 2 |
| 21 | Dijle (tidaal) | voetgangersbrug | Dijle | 104 | dag 2 |
Tabel 2-2: Een overzicht van het verloop van de estuariene meetcampagne in meetjaar 2015.
| # | Datum | Meetschip | Gemeten locaties | Databeschikbaarheid |
Opmerking |
|||
| ADCP | Diepte-staal | Troebel-heid | % | |||||
| 000 | 19/01/2015 | Pierre Petit | 7 | 7 | 7 | 7 | 100% |
Station Bazel werd nog niet aangevaren (vorig contract) |
| 20/01/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 21/01/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 001 | 16/02/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 17/02/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 18/02/2015 | Scaldis | 1 | 1 | 1 | 1 | 17% |
Campagne werd onderbroken wegens de slechte weersomstandigheden (mist) |
|
| 002 | 09/03/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 10/03/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 11/03/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 003 | 30/03/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 31/03/2015 | Scaldis | 5 | 5 | 5 | 5 | 100% |
Campagne werd onderbroken wegens de hevige wind (extra meetdag werd voorzien) |
|
| 01/04/2015 | Scaldis | 4 | 4 | 4 | 4 | 100% | ||
| 02/04/2015 | Scaldis | 4 | 4 | 4 | 4 | 100% | ||
| 004 | 20/04/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 21/04/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 22/04/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 005 | 04/05/2015 | Pierre Petit | 8 | 8 | 8 | 3 | 79% |
Geen troebelheidmetingen vanaf Antwerpen wegens een technisch probleem |
| 05/05/2015 | Scaldis | 7 | 7 | 7 | 4 | 86% |
Geen troebelheidsmetingen tussen Terhagen en Niel wegens een technisch probleem |
|
| 06/05/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 006 | 18/05/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 19/05/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 20/05/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 007 | 01/06/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 02/06/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 03/06/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 008 | 22/06/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 23/06/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 24/06/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 009 | 06/07/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 07/07/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 08/07/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 010 | 27/07/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 28/07/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 29/07/2015 | Scaldis | 6 | 6 | 6 | 0 | 67% |
Geen troebelheidsmetingen op deze meetdag wegens een technisch probleem |
|
| 011 | 10/08/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 11/08/2015 | Scaldis | 7 | 6 | 7 | 7 | 95% |
Geen ADCP-meting aan station Dijle wegens te ondiep |
|
| 12/08/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 012 | 24/08/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% |
Onbetrouwbare ADCP-data in de onderste helft |
| 25/08/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 26/08/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 013 | 07/09/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 08/09/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 09/09/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 014 | 21/09/2015 | Pierre Petit | 0 | 0 | 0 | 0 | 0% |
Meetdag opgeschort wegens onderhoud meetschip |
| 22/09/2015 | Scaldis | 9 | 9 | 9 | 9 | 100% |
Extra stations Steendorp en Temse |
|
| 23/09/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 015 | 12/10/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 13/10/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 14/10/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| 016 | 03/11/2015 | Pierre Petit | 8 | 8 | 8 | 8 | 100% | |
| 04/11/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 05/11/2015 | Scaldis | 6 | 6 | 6 | 5 | 94% |
Geen troebelheidsmetingen aan station Melle wegens technisch probleem |
|
| 017 | 07/12/2015 | Hondius | 8 | 8 | 8 | 8 | 100% | |
| 08/12/2015 | Scaldis | 7 | 7 | 7 | 7 | 100% | ||
| 09/12/2015 | Scaldis | 6 | 6 | 6 | 6 | 100% | ||
| TOTAAL | aantal | 366 | 365 | 366 | 351 | |||
| perc. (%) | 97% | 100% | 100% | 96% | ||||
2.1.2.Tijcampagnes
In een tijdsspanne van 1 week, om min of meer gelijkaardige klimatologische en tijomstandigheden te hebben, wordt op drie locaties in het estuarium een volledige tijcyclus lang de waterkwaliteit bemonsterd (Figuur 2-2). Deze campagnes worden in samenwerking met het waterbouwkundig laboratorium uitgevoerd te Kruibeke (oligohaliene zone), te Terhagen (Rumst, Rupel) en te Schoonaarde (zoete zone met korte verblijftijd). Te Lippenbroek worden jaarlijks 4 tijcycli bemonsterd, verspreid over het jaar, waarvan 1 getij bij nacht.
2.1.3.Benthische campagnes
Binnen OMES worden ook fytobenthos campagnes uitgevoerd: deze richten zich op de algen (fyto) die op de bodem (benthos) leven in de intergetijdengebieden. Staalname wordt uitgevoerd op 4 slikgebieden (Figuur 2-3), zo gekozen dat ze in de nabijheid liggen van stations waar ook de vaargeul van het estuarium wordt bemonsterd. Dit zijn de stations: Groot Buitenschoor (nabij station Grens), Boerenschans (stroomafwaarts van station Antwerpen), Ballooi (stroomopwaarts van station Temse) en Appels.
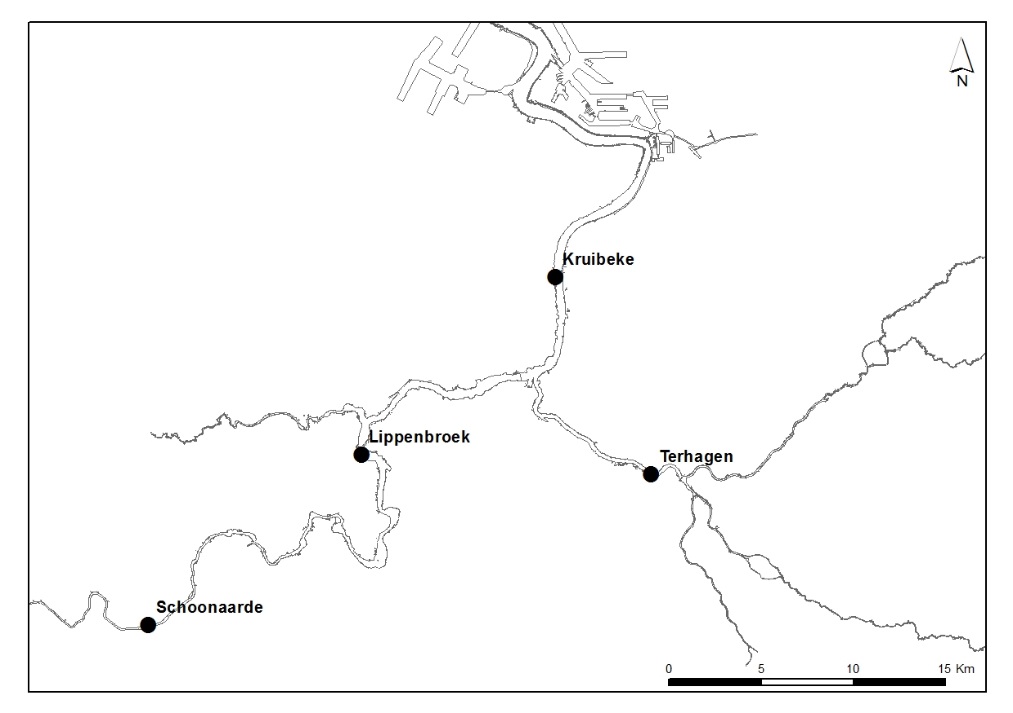
Figuur 2-2 Locaties van de estuariene tijcampagnes langs Zeeschelde en Rupel
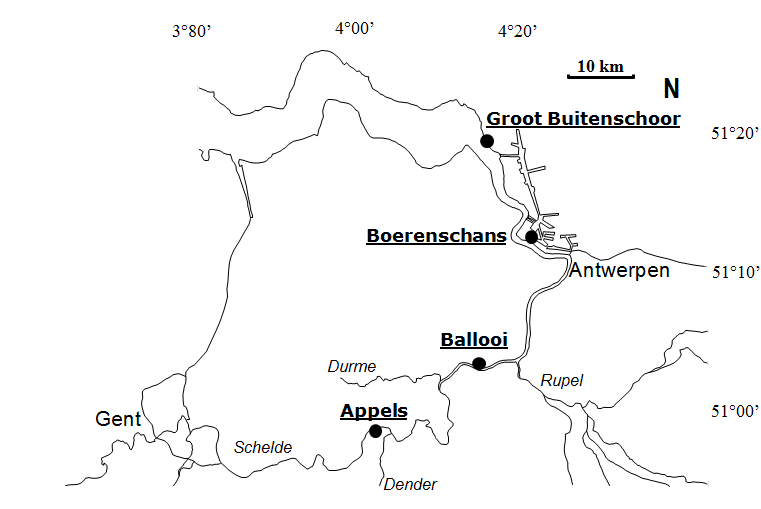
Figuur 2-3 Locaties van fytobenthische staalname langs het Schelde estuarium
2.2.Zonering
Voor een overzichtelijke rapportering van de monitoringsresultaten van alle stations, wordt vaak een zonering van het estuarium gehanteerd. Binnen MONEOS en de Evaluatiemethodiek Schelde-estuarium wordt een zonering van de Schelde voorgesteld, die ook in deze rapportage wordt aangehouden. De zonering berust op een indeling in zoutgehalte. Als maat voor het zoutgehalte worden binnen OMES de specifieke geleidbaarheid en het chloride gehalte gemeten. Op basis van het chloride gehalte werd het Schelde-estuarium ingedeeld in zones, conform de Evaluatiemethodiek (Tabel 2-3 en Figuur 2-4). De juiste ligging van deze grenzen is arbitrair: in realiteit verschuiven de chloride gehaltes onder invloed van getij en debiet, zoals besproken zal worden in hoofdstuk 3.
Tabel 2-3 Zonering in de Zeeschelde

Figuur 2-4 Zonering in de Schelde volgens zoutgehalte en verblijftijd (in de zoete zones), volgens de Evaluatiemethodiek Schelde-estuarium. Elke zone is nog verder ingedeeld in compartimenten.
2.3.Parameters en staalname
Binnen OMES worden verschillende parameters door verschillende partners gemeten (Tabel 2-4). De gebruikte analysemethoden zijn conform het OMES bestek en in overeenstemming met de vereisten binnen MONEOS. Volgende paragrafen bespreken de analysemethoden per parameter, gegroepeerd volgens de verdere indeling van het OMES rapport: Basis waterkwaliteit, Lichtklimaat en zwevende stof en Plankton. Monitoring van de parameters ten behoeve van het klimatologisch overzicht (chloride en temperatuur) wordt mee beschreven onder Basis waterkwaliteit.
2.3.1.Staalname
Voor de bepaling van de verschillende waterkwaliteitparameters worden schepstalen genomen. Stalen voor basiswaterkwaliteit, koolstofcyclus, fytoplankton en primaire productie worden allen op hetzelfde schepstaal bepaald, voor maximale compatibiliteit. Zoöplankton wordt omwille van het grote volume van het staal (50 liter) afzonderlijk, maar wel quasi gelijktijdig, bemonsterd.
De schepstalen worden genomen met een zuivere emmer. Deze wordt voorafgaand de campagne goed gereinigd en bij de eerste staalname drie maal voorgespoeld met het te bemonsteren water. Vervolgens wordt het eerste schepstaal genomen. Per locatie wordt de emmer terug voorgespoeld met het te bemonsteren water. Indien er vervuiling aanwezig is in het water (bv oliefilm) die met voorspoeling niet eenvoudig weg te spoelen valt, wordt de emmer ook tussen twee staalnames in grondig gereinigd (met detergenten). Bij het nemen van het schepstaal wordt extra aanrijking met zuurstof (door “klotsen” van het water in de emmer) vermeden.
Alle recipiënten voor stalen die verdere bepaling in het laboratorium behoeven, worden onmiddellijk na het scheppen van de emmer afgevuld, om uitzakken van bezinkbare stoffen in de emmer te vermijden. De recipiënten worden volledig afgevuld en afgesloten zonder insluiten van luchtbellen om beluchting van het staal tijdens transport te vermijden. Transport gebeurt in koelboxen met voldoende koelelementen (tot 1/4de van het koelboxvolume in de zomermaanden). Tot het moment van analyse worden de stalen zo donker en koel (5°C) bewaard, met uitzondering van de stalen voor primaire productie. Deze worden donker, maar niet gekoeld bewaard en binnen de 5 uur na staalname naar NIOZ getransporteerd.
2.3.2.Staalname voor zwevende stof
Om zwevende stof te bemonsteren worden er waterstalen van 500 tot 1000 ml genomen vanop het meetschip die nadien in een labo geanalyseerd worden op zwevende stofconcentraties. Er worden tijdens de monitoringscampagne waterstalen genomen aan het wateroppervlak (oppervlaktestalen) en in de waterkolom (dieptestalen).
Oppervlaktestalen
Voor de bepaling van zwevende stof nabij het wateroppervlak wordt 500 ml staal genomen zoals beschreven onder 2.3.1.
Dieptestalen
Sinds 2001 worden de oppervlaktestalen aangevuld met dieptestalen met behulp van een water sampler (type Van Dorn) of waterpomp. De dieptestalen worden sinds 2010 ook gebruikt om ADCP-profielen om te rekenen naar zwevende stofprofielen over de waterkolom. Hiervoor wordt er één dieptestaal in de bovenste helft van de waterkolom genomen en één dieptestaal in de onderste helft van de waterkolom. Op ondiepe meetlocaties wordt het aantal stalen beperkt tot één.
Ook tijdens dertienuurscampagnes worden frequent dieptestalen genomen met een waterpomp. Bij de dieptestalen met een pomp wordt de doorlooptijd doorheen de waterdarm steeds in acht genomen om menging van het staal met andere waterlagen te vermijden.
Tijdens het nemen van een waterstaal wordt een kabel uitgerust met een gewicht en multiparameterinstrument langzaam gevierd tot op de gewenste waterdiepte (Figuur 2-5). De waterdiepte wordt door het multiparameterinstrument geregistreerd.

Figuur 2-5: Meetopstellingen voor het nemen van dieptestalen op meetdag 1 (links) en meetdagen 2-3 (rechts) elk uitgerust met een multiparameterinstrument.
2.3.3.Samenwerking met VMM
Tijdens het zomerhalfjaar (6 maanden) wordt er twee maal per maand bemonsterd in het Schelde-estuarium. De UAntwerpen en de OMES partners nemen steeds deel aan beide campagnes, VMM enkel aan de eerste campagne. VMM meet tijdens die campagnes verschillende polluenten in de waterkolom, maar ook een aantal nutriënten. Deze nutriënten worden door VMM bemonsterd en geanalyseerd volgens de regels van de kunst (VMM beschikt over een accreditatie voor deze parameters), in overeenstemming met de vereisten uit het OMES bestek. Voor de andere parameters, en voor de zomercampagnes waaraan VMM niet deelneemt, worden de technieken gehanteerd zoals in volgend hoofdstuk vermeld.
De monitoringsresultaten van VMM (onder andere voor zware metalen en toxsiche stoffen) worden niet opgenomen in de OMES rapportage. Deze zijn wel te vinden in diverse rapportages die VMM publiceert (bijvoorbeeld voor de Kaderrichtlijn Water) en zijn te raadplegen op de VMM website. Bij de zesjaarlijkse evaluatie van het gehele Schelde-estuarium volgens de Evaluatiemethodiek Schelde-estuarium worden de OMES en VMM data voor de Zeeschelde, aangevuld met de Rijkswaterstaat gegevens voor de Westerschelde, wel samen geanalyseerd, geëvalueerd en gerapporteerd.
Tabel 2-4 Parameterlijst met vermelding van de laboratoria welke de metingen/analyses uitvoeren

2.4.Monitoring basis waterkwaliteit
2.4.1.Temperatuur, zuurstof, pH en conductiviteit
Temperatuur, zuurstofgehalte en zuurstofverzadiging worden ‘in situ’ bepaald, onmiddellijk na het afvullen van de recipiënten, rechtstreeks in de emmer. Het waterstaal werd genomen zonder aanrijking van zuurstof te veroorzaken. Meting gebeurt met een optische zuurstofmeter (Hach portable LDO) die volgens de regels van de kunst gekalibreerd werd. Meting van geleidbaarheid gebeurt met een ‘WTW LF 318’ geleidbaarheidsmeter. pH-bepaling geschiedt met een pH-meter ‘WTW pH 330’. De detectielimieten zijn weergegeven in Tabel 2-5.
Tijdens dertienuurscampagnes worden datasondes (YSI 6600 series) ingezet worden, om continu metingen doorheen de tijcyclus en diepteprofielen op te nemen van zuurstof, pH en geleidbaarheid. Deze sondes kunnen ook tijdens vaartochten ingezet worden voor diepteprofielen van bovenvernoemde parameters.
2.4.2.Chloride en sulfaat
De analyse wordt binnen de 24 uur uitgevoerd op het schepstaal.
Een 'Segmented Flow Analyser' (SFA) van het merk ‘Skalar’ wordt gebruikt voor de bepaling van chloride en sulfaat. De concentratiebepaling van chloride berust op de reactie van kwikthyocyanaat (Hg(SCN)2) met chloride-ionen. Door interactie van kwik met chloride-ionen, wordt een kwikchloridecomplex gevormd. Door behandeling met Fe3+-ionen vormt het vrijgezette kwikthyocyanaat een sterk gekleurd ijzerthiocyanaatcomplex. De absorptie van dit laatste complex wordt gemeten bij 490nm. Uit standaardcurves wordt dan de juiste concentratie bepaald.
Voor de bepaling van sulfaat wordt het staal doorheen een ionenuitwisselingskolom gepompt om kationen te verwijderen. Hierna wordt een blauw barium-methyl-thymolcomplex toegevoegd. Sulfaationen reageren hiermee en een bariumsulfaat wordt gevormd. Methyl-thymolblauw (C37H40N2Na4O13S) wordt hierbij vrijgezet. De absorptie van het methyl-thymolblauw wordt gemeten bij 460nm. Uit standaardcurves wordt dan de juiste concentratie bepaald. De detectielimieten zijn weergegeven in Tabel 2-5.
2.4.3.Stikstof (Ammonium, nitraat, nitriet en Kjeldahl-stikstof)
Een 'Segmented Flow Analyser' (SFA) van het merk ‘Skalar’ wordt gebruikt voor de bepaling van ammonium, nitriet, nitraat en na een verteringsreactie voor Kjeldahl-stikstof.
Voor de bepaling van ammonium wordt het staal na dialyse vermengd met natriumsalycyclaat (C7H5NaO3). Dit katalyseert een reactie met de vorming van een groen complex met het ammoniumion. Meting van absorptie gebeurt bij 660nm. Uit standaardcurves wordt dan de juiste concentratie bepaald.
Om een kleurreactie voor nitriet te bekomen wordt eerst een kleurreagens bereid (azijnzuur, sulfanilamide (C6H8N2O2S), α-naphtylethyleen diamine dihydrochloride (C12H6Cl2N2), gedistilleerd water). Dit vormt een complex met het nitrietion. De absorptie wordt gemeten bij 540nm. Uit standaardcurves wordt dan de juiste concentratie bepaald.
Voor de bepaling van nitraat ondergaat het staal eerst een behandeling met een ammoniumchloride buffer. Dan wordt het staal doorheen een cadmiumkolom gepompt. Door deze behandeling wordt het nitraat gereduceerd tot nitriet. Het staal wordt dan verder behandeld zoals bij nitrietbepaling. De concentratie nitraat wordt bepaald door het verschil te berekenen tussen “nitraat- en nitrietconcentratie” samen en “nitrietconcentratie”.
De bepaling van Kjeldahl-stikstof gebeurt na een vertering met zwavelzuur bij hoge temperatuur zodat alle organische stikstof wordt omgezet tot ammonium. De verdere bepaling gebeurt zoals de bepaling van ammonium.
De detectielimieten zijn weergegeven in Tabel 2-5.
2.4.4.Fosfor (Orthofosfaat en totaal fosfor)
Een 'Segmented Flow Analyser' (SFA) van het merk ‘Skalar’ wordt gebruikt voor de bepaling van orthofosfaat en na een verteringsreactie voor totaal fosfor.
Voor de bepaling van orthofosfaat wordt het staal eerst in zuur midden behandeld met ammonium-molybdaat ((NH4)6Mo7O24.4H2O). Het orthofosfaat vormt hiermee een fosfomolybdaat. Reductie van het fosfomolybdaat met ascorbinezuur resulteert in een blauw complex waarvan absorptie wordt bepaald bij 880nm. Uit standaardcurves wordt dan de juiste concentratie bepaald.
De bepaling van totaal fosfor gebeurt na een vertering met zwavelzuur bij hoge temperatuur zodat alle gebonden fosfor wordt omgezet naar orthofosfaat. De verdere bepaling gebeurt zoals de bepaling van orthofosfaat.
De detectielimieten zijn weergegeven in Tabel 2-5.
2.4.5.Silicium
Voor de Si-bepaling wordt een ‘Inductively Coupled Plasma Emission Spectrophotometer’ (ICP) van het merk ‘THERMO-OPTEC IRIS®’ gebruikt. De ICP is opgebouwd uit een kwarts toorts waardoor geïoniseerd argongas wordt bewogen. Een hoog radiofrequentieveld houdt dit gasplasma geïoniseerd. Het staal wordt ingezet en in aërosol gebracht. De hoge gasplasmatemperatuur zorgt voor een zeer goede atomisatie en ionisatie. Voor elk element resulteert dit in de emissie van een uniek lichtspectrum. Dit spectrum wordt gedecteerd door een ‘Charged Injection Device’-detector (CID-detector). De gemeten intensiteit van het SiO2 spectrum wordt gecorrigeerd op achtergrondstraling. Aan de hand van standaarden wordt de relatie bepaald tussen lichtintensiteit en silikaatconcentratie. De detectielimieten zijn weergegeven in Tabel 2-5.
Biogeen silicium (BSi) wordt bepaald na extractie met 0.1 M Na2CO3 (anhydrated) met een extractietijd van 1 uur. Analyse van Si na extractie gebeurt zoals hierboven beschreven. Vervolgens wordt een conversiefactor toegepast om de BSI concentratie te bekomen. Deze factor wordt bepaald via lineaire regressie op sequentieel geanalyseerde stalen. De sequentiële extractie gebeurt met 0.1 M Na2CO3 (anhydrated) met een extractietijd van 1, 2 en 3 uur.
Tabel 2-5 Detectielimieten voor de verschillende waterkwaliteitsparameters.
| Waterkwaliteitsparameter | Detectielimieten |
| Temperatuur | ‘WTW OXI 91’: 0°C – 40°C 'YSI 6600series': -5°C – 45°C |
| pH | ‘WTW pH 330’: 4 – 10 'YSI 6600series': 0 - 14 |
| Zuurstofgehalte | ‘WTW OXI 91’: 0 – 20 mg.L-1 O2 'YSI 6600series': 0 – 50 mg.L-1 O2 |
| Conductiviteit | 'YSI 6600series': 0 – 100 mS.cm-1 ‘WTW LF 91(318)’: 0 – 100 mS.cm-1 |
| Chloride | 1 mg.L-1 Cl- |
| Sulfaat | 4 mg.L-1 SO42- |
| Ammonium | 0,08 mg.L-1 NH4+-N |
| Nitraat | 0,05 mg.L-1 NO3--N |
| Nitriet | 0,01 mg.L-1 NO2--N |
| Kjeldahl-N | 0,30 mg.L-1 N |
| Orthofosfaat | 0,02 mg.L-1 PO43--P |
| Totaal P | 0,07 mg.L-1 P |
| Silikaat | 0,02 mg.L-1 SiO2 |
| BOD | 0,5 mg.L-1 O2 |
| Turbiditeit | 0 – 1000 NTU |
2.4.6.Biochemische zuurstofvraag BOD: CBOD en NOD
Inleiding
De biochemische zuurstofvraag (Biochemical Oxygen Demand, BOD) is de hoeveelheid zuurstof die aerobe bacteriën verbruiken in een waterstaal gedurende een bepaalde periode bij een welbepaalde temperatuur. BOD wordt meestal uitgedrukt in mg zuurstof verbruikt per liter gedurende 5 dagen incubatie in het donker bij 20°C. Hoewel op deze methode een bepaalde onzekerheid zit, wordt ze toch veel gebruikt als maat voor de hoeveelheid afbreekbaar organisch materiaal in het water.
Het zuurstofverbruik bij een BOD analyse is het resultaat van aerobe microbiële afbraak en mineralisatie van organisch materiaal of van de nitrificatie van ammonium en nitriet aanwezig in het waterstaal. Om de zuurstofvraag voor nitrificatie (Nitrogenous Oxygen Demand, NOD) uit te sluiten, wordt meestal een nitrificatie inhibitor (bijvoorbeeld Allylthioureum of Nserve) toegevoegd aan het staal. Het resultaat hiervan is dan enkel de zuurstofvraag voor koolstofmineralisatie (Carbonaceous Biochemical Oxygen Demand (CBOD))
Tabel 2-6 Overzicht van de verschillende BOD fracties
| BOD | biochemical oxygen demand |
| = CBOD + NOD | |
| CBOD | carbonacious biochemical oxygen demand |
| BOD met nitrificatie-inhibitor | |
| NOD | nitrogenous oxygen demand |
| zuurstofvraag voor nitrificatie | |
| NODth | theoretical NOD |
| = 3,22 NH4-N + 1,11 NO2-N | |
| NODmeas | measured NOD; NOD door in staal aanwezige bacteriën |
| = BODwithout - CBOD | |
| BODwithout | BOD zonder nitrificatie-inhibitor |
| = CBOD + NOD door in staal aanwezige bacteriën |
Bepaling BCOD vanaf 2010
Voor de BOD metingen in het kader van OMES, werd in de periode 1996-2009 meestal geen nitrificatie inhibitor toegevoegd (BODwithout). Jaarrond zijn er daarom niet altijd CBOD data beschikbaar voor 2009. Na 2009 werd standaard Allylthioureum toegevoegd als nitrificatie inhibitor. Vanaf 2010 zijn hierdoor jaarrond CBOD data beschikbaar voor alle stations (zie Figuur 2-6 voor een overzicht).

Figuur 2-6 Overzicht van de meetperioden van de verschillende BOD fracties. In de vol gekleurde perioden werd de fractie elke campagne bepaald. In de gearceerde periode zijn de data gedeeltelijk aanwezig.
De analyse wordt binnen de 24 uur ingezet op het schepstaal. Het watermonster wordt verdund met zuurstofrijk verdunningswater en geënt met commercieel beschikbaar entmateriaal (micro-organismen). De nitrificatie wordt onderdrukt door toevoegen van een inhibitor: allylthioureum (ATU). Deze verdunde monsters worden gedurende 5 dagen ± 6u geïncubeerd bij (20 ± 2)°C in het donker in een volledig gevulde en afgesloten Winkler-fles. Voor en na de incubatie wordt het zuurstofgehalte gemeten. Vervolgens kan hieruit het zuurstofverbruik door biochemische oxidatie berekend worden.
De BOD bepaling dient binnen de 24 uur gestart te worden. Als alternatief kunnen de verdunde monsters eerst gedurende 48u ± 2uur bewaard worden bij een temperatuur van 1°C-5°C en vervolgens gedurende 5 dagen geïncubeerd worden bij een temperatuur van 20°C ± 2°C.
De detectielimieten zijn weergegeven in Tabel 2.1.
Bepaling langetermijntrends CBOD en NOD voor 2009
-
Zoete zones
Meestal werd voor 2009 geen nitrificatie inhibitor toegevoegd. Om toch het onderscheid te maken tussen CBOD en NOD werden in de periode 1998-2003 verschillende BOD analyses parallel met en zonder nitrificatieremmer (Nserve) uitgevoerd. Op basis van deze metingen wordt NODmeas bepaald:
NODmeas = BODwithout – CBOD
De theoretische zuurstofvraag voor nitrificatie (NODth) kan ook stoichiometrisch bepaald worden, gebaseerd op de hoeveelheid zuurstof nodig om ammonium en nitriet om te zetten tot nitraat:
2 NH4+ + 3 O2 ->2 NO2- + 2 H2O + 4 H+ + energie
2 NO2- + O2 -> 2 NO3- + energie
Omdat nitrificerende bacteriën ook een deel van hun zuurstofbehoefte zullen halen uit de reductie van CO2 bij lithotrofe biosynthese, worden beter de experimenteel afgeleide waarden van Wezernak & Gannon (1967) gebruikt om de zuurstofvraag voor nitrificatie te berekenen in plaats van de stoichiometrische ratio’s. Zij vonden de zuurstof-stikstof ratio bij anorganische stikstof oxidatie gelijk was aan 3.22 mg/l zuurstof per mg/l ammonium-stikstof omgezet tot nitriet en 1.11 mg/l zuurstof per mg/l nitriet-stikstof omgezet tot nitraat.
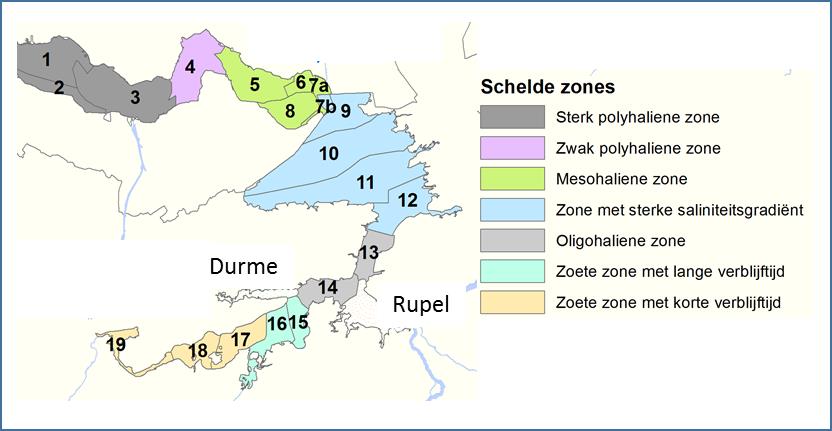
Het nitrificatieproces gedurende een BOD meting is echter afhankelijk van de aanwezigheid van een actieve populatie van nitrificerende bacteriën. Omdat bij de BOD experimenten geen inoculum van nitrificerende bacteriën werd toegevoegd, was het nitrificatieproces dus afhankelijk van de bacteriële populaties op dat moment aanwezig in het estuarium. Wanneer echter de watertemperatuur daalt onder de 15°C, is de aanwezigheid van een actieve populatie nitrificerende bacteriën niet steeds verzekerd, wat leidt tot grotere verschillen tussen de opgemeten NODmeas en de berekende NODth (Figuur 2-7). Voor watertemperaturen boven 15°C bestaat wel een mooie correlatie tussen NODmeas en NODth.
Voor de bepaling van langetermijn trends kunnen we zo beschikken over data van de biochemische zuurstofvraag voor zowel nitrificatie als koolstofmineralisatie, voor de gehele OMES periode, voor watertemperaturen boven 15°C, voor de meeste stations in het zoetwatergetijdengebied. Voor de gehele OMES periode werden de NOD waarden berekend (=NODth). Voor de periode 1996-2009 werd de CBOD berekend (CBOD = BODwithout –NODth); vanaf 2010 werd CBOD rechtstreeks gemeten.
Figuur 2-7 a. Relatieve afwijking van NODmeas ((NODmeas – NODth)/NODmeas) in functie van de watertemperatuur. b. Correlatie tussen NODth en NODmeas aan het station Dendermonde voor watertemperaturen boven 15°C.
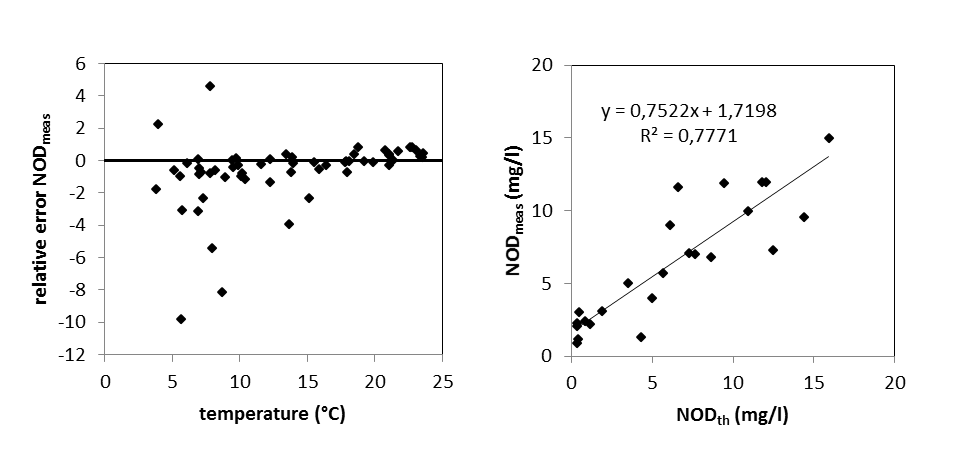
Aan de ‘boundaries’ Bovenschelde en Dender wordt geen goede correlatie gevonden tussen NODmeas en NODth. Hoewel in de zijrivieren hoge concentraties voorkwamen van ammonium of nitriet, welke hoge NODth opleverden van soms meer dan 10 mg/l, werden tussen de BOD metingen met en zonder nitrificatie inhibitor (CBOD en BODwithou) amper verschillen opgemeten (Figuur 2-8 voor Bovenschelde). Dit wijst erop dat er weinig tot geen nitrificatie-activiteit aanwezig is in de zijrivieren. Blijkbaar is er daar geen actieve populatie nitrificerende bacteriën, zelfs niet bij temperaturen boven 15°C. Daarom zijn voor de periode dat er enkel BODwithout data beschikbaar zijn, deze data verder beschouwd als CBOD data. De NOD die voor deze station berekend werd, stelt in dit geval de potentiële zuurstofvraag voor nitrificatie voor, indien nitrificerende bacteriën aanwezig zouden zijn.
Figuur 2-8 correlatie tussen CBOD en BODwithout in de Bovenschelde, bij temperaturen boven 15°C
Aan het meest opwaartse station Melle komen de NODth en de NODmeas data soms overeen, maar vaak ook niet, wat erop wijst dat zo kort bij de boundary van het estuarium er niet steeds een actieve populatie nitrificerende bacteriën aanwezig is. Daarom konden geen CBOD en NOD data bepaald worden voor de ganse OMES periode voor dit station.
-
Oligohaliene zone en zone met sterke saliniteitsgradiënt
In de oligohaliene zone en de zone met sterke saliniteitsgradiënt konden geen betrouwbare correlaties gevonden worden tussen NODmeas en NODth, wellicht omwille van lagere concentraties aan CBOD en NOD (zie Resultaten) en dus grotere relatieve fout.
-
Rupel
Ook voor het station Boom op de Rupel werd de theoretisch berekende NODth (op basis van NH4 en NO2) vergeleken met de op metingen van BODwithout en CBOD gebaseerde NODmeas. In de Rupel blijken deze niet gecorreleerd te zijn (zie Resultaten), ook niet bij temperaturen boven 15°C. Voor de Rupel zijn er bijgevolg geen CBOD data beschikbaar voor 2009.
2.4.7.Opgelost organisch koolstof (Dissolved Organic Carbon, DOC)
De staalname van DOC gebeurt door water te filtreren met een glazen spuit door een glasvezel filter (type Whatmann GF/F, 0.7 µm) in een glazen proefbuis. Al het glaswerk, inclusief de glasvezel filter worden vooraf verhit, minimum 8 uur op 450°C om al het organisch materiaal te verwijderen. De DOC concentratie wordt colorimetrisch bepaald (Skalar).
2.4.8.Opgelost anorganisch koolstof (Dissolved Inorganic Carbon, DIC): concentratie en isotoop verhouding (δ13C-DIC)
Voor de bepaling van DIC concentraties zijn 2 belangrijke parameters vereist namelijk pH en totale alkaliniteit (TALK), alsook temperatuur en saliniteit van het water. De pH, temperatuur en saliniteit van de waterstalen wordt gemeten bij de staalname. Voor de bepaling van TALK worden 50 ml waterstalen gefilterd (GFF, 0.7 µm) en gefixeerd met kwikchloride en bewaard bij 4°C. De analyse gebeurt binnen de maand na staalname. De totale alkaliniteit wordt bepaald met een automatische titrator (Mettler-Toledo, precisie 0.3%) door een bepaald volume staal met HCl van gekende normaliteit (Titrisol standaard 0.01N) te titreren tot het eindpunt voor bicarbonaat. Totale anorganische koolstof, bicarbonaat, carbonaat en CO2 concentraties worden dan berekend vanaf TALK en pH, temperatuur en saliniteit via de “CO2SYS” Excel macro van Lewis and Wallace (2006).
Voor de bepaling van de isotopenratio van DIC (δ13C-DIC), wordt 10 ml staal luchtledig bewaard op 4°C na fixatie met kwikchoride. De analyse gebeurt vervolgens binnen de maand met een Isotopic Ratio Mass Spectrometer (IRMS) via een gas-bench sampler (Gasprep-Nu perspective IRMS, Nu Instruments).
2.4.9.Particulair organische koolstof (Particulate Organic Carbon, POC) en koolstof/stikstof (C/N) verhouding
De hoeveelheid particulair organisch koolstof aanwezig in de waterkolom wordt bepaald door katalytische combustie van dit materiaal verzameld op een glasvezel filter van 0.7µm nominale porositeit (type Whatmann GF/F). Deze filters werden vooraf gedecontamineerd door ze gedurende 8u te verwarmen op 450°C. Een zo groot mogelijk volume water wordt gefiltreerd, dit varieert meestal van 200 ml tot 500 ml afhankelijk van de turbiditeit.
Filters wordt vervolgens gedurende 2 uur gedroogd bij 105°C zoals bepaald in de standaardprocedure van het Belgisch instituut voor de normalisatie. Blank stalen (i.e. gedecontamineerde filters die de hele procedure doorlopen hebben met uitzondering van de filtratie) worden op regelmatige tijdstippen geanalyseerd.
Voor de analyse wordt een filterstukje met gekend oppervlak uitgesneden en vervolgens gedurende 8 uur in een verzadigde HCl atmosfeer geplaatst ten einde de carbonaat te verwijderen (CO2 vrijzetting). Vervolgens wordt dit filterstukje in een zuiver tincupje geborgen, waarna de eigenlijke analyse volgt met een Elemental Analyser (Thermo-Finnigan Flash EA 1112). Bij deze wordt het organisch materiaal verbrand bij 1010°C onder aanvoer van zuivere O2, waarna overmaat O2 en NOx gassen gereduceerd worden op metallisch koper bij 600°C. Daarna wordt de waterdamp weggevangen en worden CO2 en N2 chromatografisch gescheiden en bepaald met behulp van een TCD (Thermal Conductivity Detector). Kalibratie gebeurt met nauwkeurig afgewogen acetanilide (C8H9NO) hoeveelheden. De gemeten koolstof is de particulair organische koolstof en de stikstof is de totale particulaire stikstof (in hoofdzaak = organische N). Op basis van gemeten POC als PN gehalte, wordt vervolgens de C/N verhouding berekend. De precisie voor de analyse van particulair organische koolstof en stikstof is respectievelijk 5% en 4%.
Naast de metingen van POC conform bestek, zullen er ook extra metingen gebeuren voor de 13C isotoop verhouding van POC (δ13C-POC) door middel van een EA-IRMS systeem (elemental analyzer – isotope rtio mass spectrometer).
2.4.10.Particulair totaal en anorganisch koolstof (TOC en PIC)
Voor de bepaling van totaal particulair koolstof en particulair anorganische koolstof wordt dezelfde procedure toegepast als voor POC. Een tweede uitgesneden stukje filter ondergaat echter geen eliminatie van de aanwezige carbonaten. Op deze manier wordt de totale hoeveelheid koolstof bepaald (TOC). De hoeveelheid anorganische koolstof wordt bijgevolg berekend door het verschil te nemen tussen de totale koolstofhoeveelheid en de organische koolstofhoeveelheid (PIC = TOC-POC). Resultaten hebben tot nu toe aangetoond dat TOC en POC niet significant verschillen waardoor PIC verwaarloosbare concentraties vertonen. PIC wordt daarom niet verder besproken.
2.5.Monitoring lichtklimaat en zwevende stof
2.5.1.Lichtextinctiecoëfficiënt en eufotische diepte.
Voor het bepalen van het lichtklimaat wordt de lichtextinctiecoëfficiënt kD gemeten: de mate waarin licht afneemt met de diepte, door de troebelheid van het water. Hiertoe worden twee lichtsensoren (PAR sensor LiCOR) gebruikt die op een gekende afstand van elkaar (Δz = 40 cm) worden gemonteerd. Het invallende licht kan nu gelijktijdig op twee verschillende dieptes gemeten worden. Voor een meting moeten geen sensoren op en neer bewogen worden, en fluctuaties in het invallend licht vormen geen probleem: ze worden door beide sensoren gelijktijdig geregistreerd. Wanneer men uitgaat van een exponentiële afname met de diepte van het invallende licht, kan de extinctiecoëfficiënt via volgende formule bepaald worden:
![]()
waarbij E1 de lichtinstraling aan de bovenste sensor is, E2 de instraling aan de onderste sensor en (Δz) de verticale afstand tussen beide sensoren. Het is dus mogelijk om kD continu te meten, zelfs bij sterk fluctuerende lichtinstraling.
Aan elk estuarien monitoringsstation wordt kD bepaald. Hiertoe wordt de opstelling met twee lichtsensoren gedurende 3 tot 5 minuten net onder het wateroppervlak gehangen. Elke seconde wordt de aanwezige lichtintensiteit en kD bepaald, elke 5 seconden wordt een gemiddelde waarde voor deze parameters geregistreerd. Achteraf worden outliers verwijderd: ten gevolge van deining en golfslag kan de bovenste lichtsensor soms boven water komen, met foutieve waarden tot gevolg. Vervolgens wordt voor elk station een gemiddelde kD (met standaard deviatie) berekend.
Vanuit kD kan de eufotische diepte (Zeu) worden bepaald. Zeu is de diepte tot waar licht kan doordringen in de waterkolom, gedefinieerd als de diepte waar de hoeveelheid invallend licht nog 1% bedraagt van deze aan het wateroppervlak. Zeu kan als volgt berekend worden: Zeu = -1/kD * ln (0.01) ≈ 4,6/ kD.
2.5.2.Zwevende stof
Het waterstaal met gekend volume wordt gefiltreerd op een filter van gekend gewicht (tarra). Voor alle waterstalen wordt de filter nagespoeld met 3x50 ml gedestilleerd water volgens de norm NBN-EN872 (1996). Het monster wordt gedroogd en na afkoeling opnieuw gewogen. De zwevende stof wordt bepaald door het verschil van totaal gewicht en tarra te berekenen.
2.5.3.Troebelheidsmetingen (turbiditeitsmetingen)
In functie van het lichtklimaat worden systematisch troebelheidsmetingen geregistreerd via een multiparameterinstrument (van RBR type XR-420 CTD+ of gelijkaardig). Het instrument wordt samen met de water sampler gevierd in de waterkolom (Figuur 2-5). Tijdens het profileren wordt er minstens elke 10 seconden een troebelheidswaarde geregistreerd.
De troebelheidssensor wordt vooraf geijkt in het labo met standaard formazineoplossingen zodanig dat er telkens een gestandaardiseerde waarde wordt geregistreerd over de verschillende meetcampagnes. Bovendien geven deze troebelheidsmetingen een maat voor zwevende stofconcentraties na correlatie met de genomen waterstalen. Dit vormt een aanvulling op de waterstalen en ADCP-profielen.
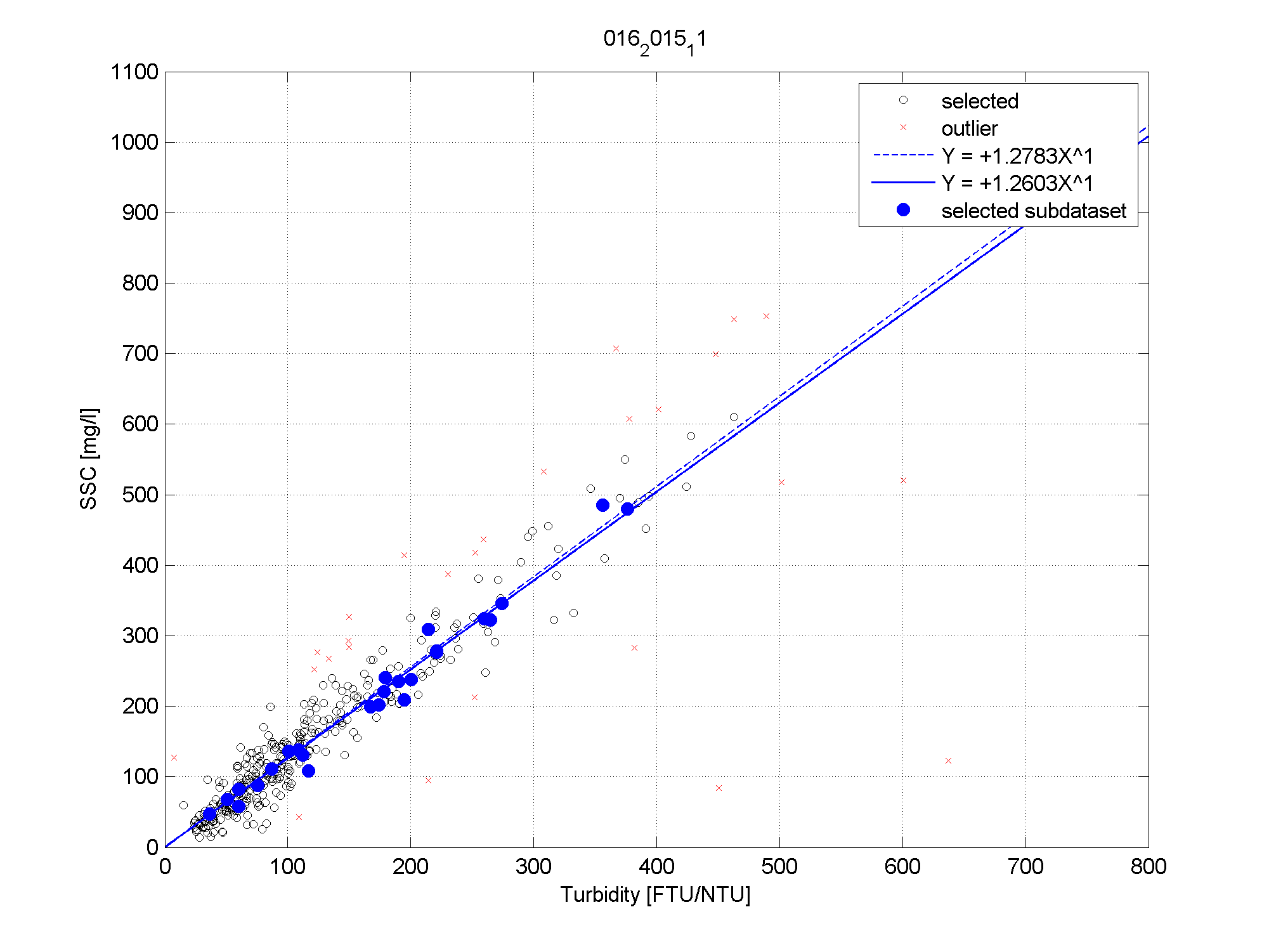
Figuur 2-9: De correlatie tussen troebelheid (X-as) en zwevend stofconcentratie (Y-as) voor het volledige meetjaar 2015 (zwarte punten en correlatie, y = 1.2783x) en meetcampagne 11-2015 (blauwe volle punten en correlatie, y = 1.2603x).
Op basis van iedere meetcampagne wordt telkens de correlatie tussen zwevende stof (afkomstig van de dieptestalen) en troebelheid (gemeten door de multiparameterinstrument) bepaald. Figuur 2-9 toont een voorbeeld van zo’n correlatie voor de meetcampagne 11-2015 en voor het volledige meetjaar 2015. Beide correlaties vertonen een duidelijk verband tussen troebelheid en zwevende stof.
2.5.4.zwevende stof en stroomsnelheid via ADCP
meetmethode
Vanaf 2011 worden de meetschepen uitgerust met een ADCP (van Teledyne RDI) en externe dGPS (van Hemisphere) om de stroomsnelheden, stroomrichtingen en onrechtstreeks zwevende stofconcentraties over de waterkolom te bepalen. Tijdens het nemen van de dieptestalen op een meetlocatie (punt) wordt er continu gemeten met een ADCP (gedurende +/- 5 minuten) waarvoor voor iedere meetcel van 0.25 of 0.50m meetwaarden wordt geregistreerd. De verticale celgrootte van 0.50m wordt gehanteerd op de meest afwaartse en diepere meetlocaties, en 0.25m voor de opwaartse en ondiepere meetlocaties.

Figuur 2-10: De ADCP meetopstelling op meetschip Scaldis I (meetdagen 2-3).
Via deze akoestische meettechniek worden stroomsnelheden berekend uit het verschil tussen totaal gemeten snelheid en de geregistreerde vaarsnelheid. De vaarsnelheid wordt door de ADCP geregistreerd door een ingebouwde bottom-trackmodule die tevens de bodemligging vaststelt. Omwille van mogelijke magnetische interferentie tussen het meetschip en het intern kompas van het instrument wordt het meetschip uitgerust met een dGPS. Diepte-, temperatuur- en geleidbaarheidsprofielen worden simultaan met de dieptestalen genomen om het akoestische achtergrondsignaal (zie Figuur 2-5) te corrigeren en de diepteligging van de cellen goed te kunnen positioneren in de waterkolom. De 3 parameters worden geregistreerd door een multiparameterinstrument (van RBR type XR-420 CTD+ of gelijkaardig) en omgerekend naar saliniteit op basis van Practical Salinity Scale van 1978 (UNESCO, 1983).
Om de ruimtelijke variatie van zwevende stof rondom een station of meetpunt te karakteriseren, worden er op bepaalde stations vaste raaien gevaren (Tabel 2-7, Figuur 2-1). Tijdens het varen wordt er met de ADCP continu gemeten zodanig niet enkele een verticaal profiel voor stromingen en zwevende stof aanwezig is maar ook voor heel de dwarssectie over de desbetreffende waterloop. Deze methodiek wordt ook toegepast aan GGG Lippenbroek tijdens de 24-uursmetingen.
Tabel 2-7: Een overzicht van meetstations waar ADCP-raaien gevaren wordt, inclusief de coördinaten van de theoretische raaien (in UTM31, WGS84) (zie ook Figuur 2-1).
| id | station | X rechteroever [m] | Y rechteroever [m] | X linkeroever [m] | Y linkeroever [m] |
| 1 | Grens | 587320 | 5689874 | 586388 | 5688916 |
| 2 | Liefkenshoek | 590573 | 5684054 | 590242 | 5683184 |
| 4 | Antwerpen | 597597 | 5675675 | 597222 | 5675802 |
| 5 | Kruibeke | 593024 | 5670309 | 592605 | 5670309 |
| 7 | Temse | 585579 | 5663806 | 585626 | 5664135 |
| 8 | Lippenbroek | 582387 | 5660350 | 582232 | 5660148 |
| 10 | Dendermonde | 577812 | 5654509 | 577728 | 5654615 |
| 13 | Uitbergen | 567438 | 5651875 | 567350 | 5651887 |
| 18 | Terhagen | 598316 | 5659259 | 598429 | 5659150 |
Verwerking van ADCP data tot zwevende stof
Met behulp van de software Sediview, wordt de verzamelde ADCP data verwerkt. Tijdens het verwerkingsproces wordt:
- De diepteligging van de meetcellen gecorrigeerd op basis van het verzameld saliniteitsprofiel;
- Het akoestisch achtergrondsignaal gestandaardiseerd en omgerekend naar zwevende stofconcentraties op basis van de verzamelde dieptestalen en troebelheidsmetingen.
De theorie achter de omzetting van het ADCP achtergrondsignaal naar zwevende stof is complex. De theorie wordt gebundeld in de software Sediview en worden parameters S, Ks, αw en αs bepaald, gebruikmakend van volgende vereenvoudigde relatie (DRL, 2003):
![]()
M(r) = massaconcentratie per eenheidsvolume op diepte (r);
S = relatieve backscattercoëfficiënt;
Ks = omgevings- en instrumentconstante;
dB = de gemeten relatieve akoestische achtergrondintensiteit;
αw = waterabsorptiecoëfficiënt;
αs = sedimentabsorptiecoëfficiënt (in functie van de sedimenteigenschappen).
De omzettingsparameters variëren per station, ADCP-type en tijd waardoor deze bij elke campagne en station opnieuw bepaald en doorgerekend dienen te worden. Dit gebeurt telkens op gekende kalibratiemomenten waar een dieptestaal genomen wordt. Een dieptestaal zal steeds genomen worden in het meetgebied van de ADCP: onder de insteekdiepte van de ADCP en 1 à 2m boven de interferentiezone (Figuur 2-12). Troebelheidsmetingen omgerekend naar zwevende stofconcentraties worden als een extra kalibratiepunt meegenomen tijdens het verwerkingsproces om de onzekerheid op berekende zwevende stofprofielen te verlagen. Het eindresultaat wordt vervolgens gecontroleerd in onderstaande correlatiegrafieken (Figuur 2-11).
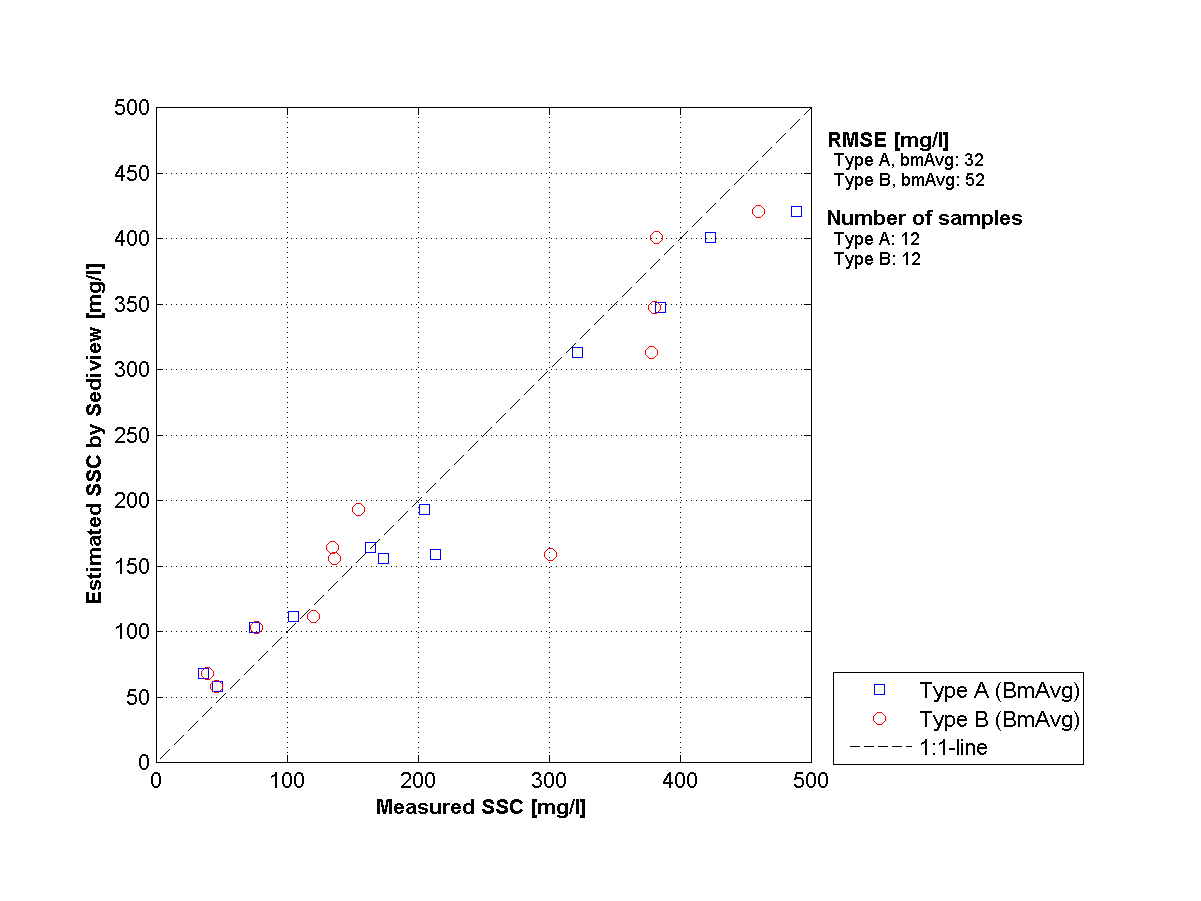
Figuur 2-11: Een voorbeeld van een correlatie tussen geschatte zwevende stof na Sediviewkalibratie (ADCP) in functie van zwevende stof afkomstig van de waterstalen (type A) en troebelheid (type B).
Het bekomen resultaat uit Sediview wordt vervolgens gefilterd op uitschieters. De stationaire ADCP-metingen worden omgerekend naar een verticaal profiel door het gemiddelde te berekenen over de tijd. Voor de raaimetingen worden de verwijderde uitschieters opgevuld door interpolatie. Na dataprojectie op de theoretische raai wordt het waterdebiet, de sedimentflux en de gemiddelde zwevende stofconcentratie afgeleid uit de raaimetingen. Hierbij worden de niet-gemeten zones geschat door een berekende waarde via extrapolaties. De niet-gemeten zones zijn een gevolg van onbereikbare plaatsen voor het meetschip (aan de oevers), de insteekdiepte van de ADCP en akoestische interferentie aan de waterbodem (Figuur 2-12). De niet-gemeten zone nabij de bodem bedraagt ongeveer 6% van de waterdiepte.
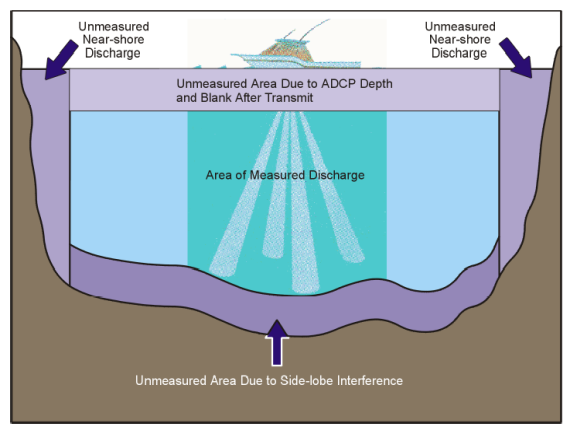
Figuur 2-12: Aanduiding van de niet-gemeten zones tijdens het uitvoeren van een ADCP-raai (RD Instruments, 2001)
De zone aan de het wateroppervlak wordt geschat op basis van de gemiddelde snelheid of zwevende stofconcentratie van de 3 onderliggende cellen. Voor de zone nabij de bodem wordt de zwevende stofconcentratie geschat op basis van de gemiddelde concentraties van de onderste 5 gemeten cellen die verhoogd wordt met een factor 1,25. De stroomsnelheden nabij de bodem worden geschat via Chen’s machtsvergelijking:
![]()
z = afstand tot de waterbodem [m]
u = snelheid op afstand z [m/s]
u* = wrijvingssnelheid [m/s]
z0 = ruwheidshoogte [m]
b = Exponent (1/6)
De niet-gemeten zones nabij de oevers wordt voor zowel water- als voor sedimentdebiet geschat op basis van het gemiddelde van de 5 dichtstbijzijnde profielen en schatting van het niet-gemeten oppervlak in de waterkolom. Het oppervlak wordt berekend op basis van de afstand oever tot dichtstbijzijnde ADCP-profiel, diepte en correctiefactor voor helling van de oever (0,35).
2.5.5.Data-analyse van zwevende stof tot jaargemiddelden
Homogenisatie van de data
Binnen OMES is gekozen om tijonafhankelijk te bemonsteren. Dit impliceert dat de staalname op verschillende momenten in de tijcyclus wordt uitgevoerd, met onvermijdelijk een zekere variatie in zwevende stof tot gevolg. Door uit te middelen over seizoenen of jaren, kan ook de tijvariatie grotendeels uitgevlakt worden. Deze gemiddeldes zullen gebruikt worden voor het uitvoeren van een analyse van zwevende stof over de jaren heen en de ruimtelijke variatie in het meetgebied.
Omdat er tijdens bepaalde jaren meer metingen zijn uitgevoerd tijdens de zomermaanden dan tijdens de wintermaanden wordt van de volledige dataset eerst een daggemiddelde en vervolgens een maandgemiddelde van de metingen genomen. Daarna worden jaargemiddeldes (januari-december), zomergemiddeldes (juni-november) en wintergemiddeldes (december-mei) berekend op basis van de maandgemiddeldes.
Om een bias te vermijden omdat sommige meetlocaties dichter bij elkaar liggen dan andere, worden de data gehomogeniseerd in de ruimte. Hiervoor wordt de Zeeschelde verdeeld in secties met een lengte van 10 km, en wordt uitgemiddeld over elke sectie. Tenslotte worden systeemgemiddelden berekend voor verschillende zones, namelijk de Beneden-Zeeschelde, de Boven-Zeeschelde, de zone tussen 100 en 130 km van de monding (ruwweg tussen Lippenbroek en Appels) en tussen 60-80 km van de monding (ruwweg de zone rond Kallo-Antwerpen).
Analysemethodiek van de dieptestalen en ADCP-data
Zowel aan het wateroppervlak als dieper in de waterkolom worden er stalen genomen tijdens de monitoringscampagne. De dataset van de oppervlaktestalen is het meest uitgebreid in de tijd en is reeds gestart in 1995. Het nemen van dieptestalen is later begonnen in 2001, maar verdient ook aandacht omdat deze bijkomende informatie kan leveren over de verticale verdeling van de sedimentconcentratie, en kan dienen als (quasi-)onafhankelijke controle van de analyse van de oppervlaktestalen.
Over de jaren heen werden er verschillende meetprotocollen gehanteerd voor het bemonsteren van de stalen waardoor de diepteligging van de stalen en het aantal stalen varieert over de tijd (Figuur 2-13). Tussen 2001 en 2004 werden de dieptestalen bemonsterd op willekeurige dieptes en in de periode 2005-2009 werd er meer bemonsterd op vaste percentages in de waterkolom (10%, 25%, 50%, 75%, 90%). Sinds 2012 worden de dieptestalen voornamelijk genomen in functie van de ADCP-metingen, waardoor er minder stalen worden genomen in de onderste en bovenste 20% van de waterkolom. Deze zones worden minder bemonsterd omdat deze zones niet bemeten worden door de ADCP.
Om de mogelijkheid op een bias te vermijden wordt in de analyse enkel gewerkt met stalen die genomen zijn tussen 20% en 80% van de waterdiepte. Daarnaast zijn er in 2001 slechts een klein aantal stalen genomen, wat niet voldoende is om tot een representatief jaar- of seizoensgemiddelde te komen. De metingen uit 2001 worden daarom niet beschouwd; enkel metingen uit de periode 2002-2015 worden verder beschouwd in de analyse.
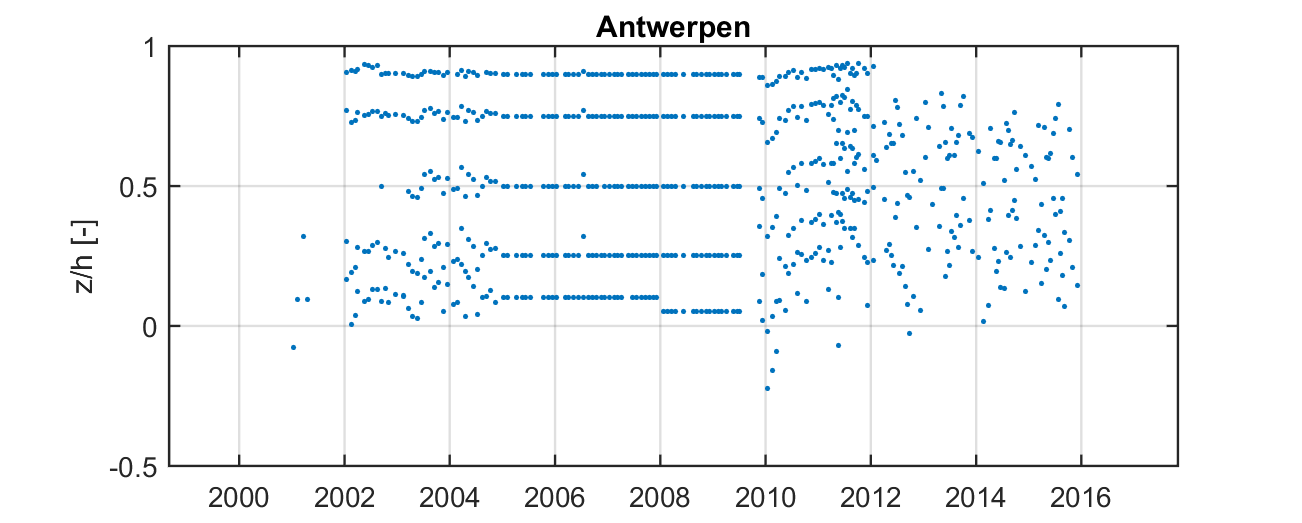
Figuur 2-13: Verdeling van dieptestalen over de waterkolom doorheen de tijd op meetlocatie Antwerpen. De verticale as geeft de relatieve afstand boven de bodem weer (z/h=0 op de bodem, z/h=1 aan het wateroppervlak).
Om de volledige dataset van de dieptestalen onderling te kunnen vergelijken werd voor elke meting de relatieve afstand boven de bodem z/h bepaald, waarbij hde lokale en instantane waterdiepte voorstelt (z/h=0 op de bodem, z/h=1 aan het wateroppervlak). Om dit te doen is voor elke meting het lokale waterpeil en bodempeil nodig op het moment van de meting.
Het waterpeil (in m TAW) werd bepaald uit de naburige getijmeetstations. In de ADCP-metingen uit de periode 2011-2015 werd steeds ook de bodemdiepte op de staalnamepunten gemeten met behulp van de ADCP-bottom tracker. In de databestanden wordt het bodempeil soms weergeven als absoluut peil (in m TAW) of als diepte ten opzichte van het wateroppervlak (in m). In het tweede geval werd de bodemdiepte omgezet naar een absoluut bodempeil (in m TAW) met behulp van de geregistreerde waterdiepte in de nabijgelegen getijstations op het moment van de staalname. Voor de jaren voor 2011 is de bodemdiepte op de meetlocaties niet exact gekend. Daarom wordt voor alle metingen die zijn uitgevoerd voor 2011 het gemiddelde gemeten bodempeil uit de jaren 2011 en 2012 aangenomen als bodempeil. In de Beneden-Zeeschelde (tot ca. Antwerpen) zijn verdiepingen van de Schelde uitgevoerd in 1970, 1995 en 2010; deze worden hier dus niet in rekening gebracht omdat er geen dieptemetingen beschikbaar waren op de locaties van de staalnames.
Nadat de volledige tijdreeks was opgesteld van het bodempeil en het waterpeil op elke meetlocatie, werd voor elk staal de relatieve afstand boven de bodem bepaald. Het is duidelijk dat er een onzekerheid is over zowel het waterpeil als het bodempeil op het moment van de staalname, wat zorgt voor een onzekerheid op de relatieve diepte van elk genomen staal.
Vooraleer dat de dataset werd gehomogeniseerd, werd deze onderworpen aan een visuele kwaliteitscontrole. Uitschieters, gedefinieerd als metingen die meer dan tien maal de interkwartielafstand boven de 75ste percentiel liggen van alle metingen op een bepaalde meetlocatie, werden verwijderd.
Sinds 2011 wordt zwevende stof in de waterkolom ook bepaald aan de hand van ADCP-metingen. De dataset van ADCP-metingen zijn geanalyseerd op dezelfde manier als de dataset van de eigenlijke dieptestalen.
In Hoofdstuk 5 wordt er aangetoond dat er een correlatie bestaat tussen troebelheid en zwevende stof. Gezien dat er sinds 2015 ook troebelheidsdata wordt verzameld over de diepte, kunnen deze profielen bijkomende informatie geven over zwevende stof over de diepte. De bijkomende optische meettechniek kan vergeleken worden met de dieptestalen en akoestische meettechniek (ADCP-profielen). Figuur 2-14 en Figuur 2-15 illustreren een vergelijking van de verschillende meettechnieken. Bij lage concentraties (kleiner dan ca. 200 mg/l) vertonen de meettechnieken een gelijkaardig gedrag en zijn de dieptegemiddelde data van dezelfde grootteorde. De variatie tussen de verschillende technieken neemt toe met de sedimentconcentratie (Figuur 2-13). Een mogelijke verklaring hiervoor is dat bij hoge concentraties het gehalte aan zwevende stof in de waterkolom zeer variabel is in tijd. Zo worden de dieptestalen genomen op een bepaald moment in de tijd terwijl de ADCP-profielen, en in mindere mate de troebelheidsprofielen, een gemiddelde zijn over de tijd. Dit kan een verschil in de diepteprofielen geven tussen de verschillende meettechnieken. In Figuur 2-13 is het jaargemiddelde van meetjaar 2015 tussen beide meettechnieken vergeleken waar de gemiddelde afwijking +/- 20 mg/l bedraagt.
Daarenboven zijn de ADCP- en de troebelheidsmetingen 2 verschillende meettechnieken (resp. akoestisch en optisch) die gevoelig zijn voor bepaalde en andere randeffecten. De verwerkings- en omzettingsmethode naar zwevende stofconcentraties zullen hierdoor ook verschillen (zoals hierboven beschreven). Zo dooft bijvoorbeeld het akoestisch signaal uit in de diepte ten gevolge van water- en sedimentabsorptie. In de Sediview-software wordt deze signaaluitdoving gecompenseerd maar bij hoge concentraties (> 400 mg/l) en in de brakwaterzone kan het zijn dat deze onvoldoende gecompenseerd wordt. Hierdoor kunnen de concentraties artificieel afnemen over de diepte. In Figuur 2-14 wordt een extreem voorbeeld geïllustreerd aan meetlocatie Antwerpen tijdens meetcampagne 12-2015.
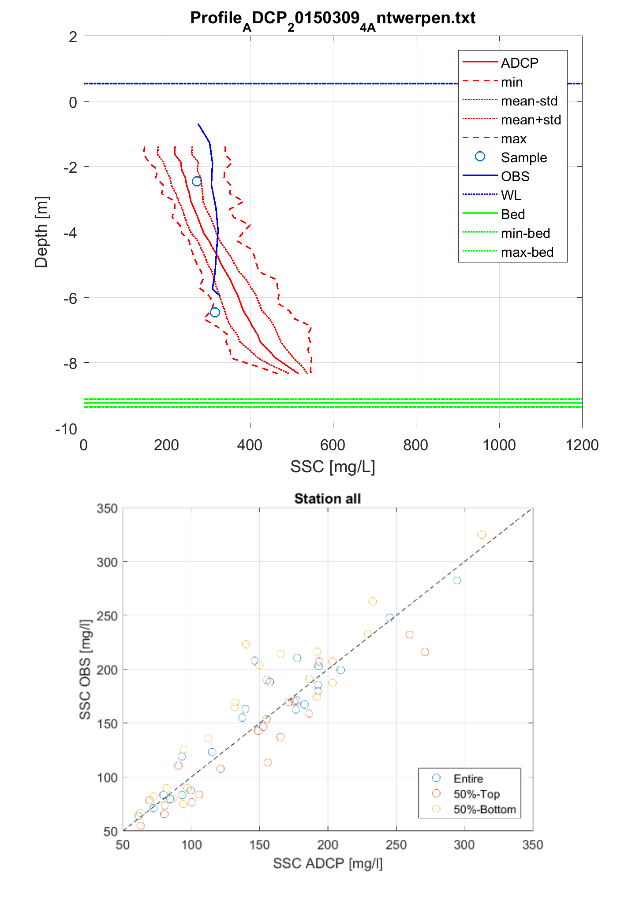
Figuur 2-14: Diepteprofielen van zwevende stof op basis van staalname, ADCP (akoestisch) en troebelheid (optisch) aan station Antwerpen (boven) voor meetcampagne 03-2015, met aanduiding van de variatie rondom een ADCP-profiel. Onder: een cross-plot tussen de zwevende stofprofielen van troebelheid (OBS) en ADCP voor jaargemiddelde over een volledige profiel (entire), bovenste helft (50%-top) en onderste helft (50%-onder) voor alle stations.
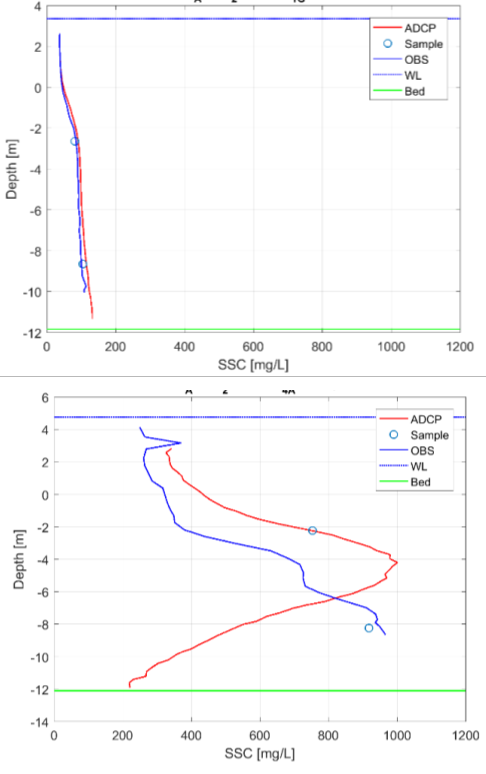
Figuur 2-15: Diepteprofielen van zwevende stof op basis van staalname, ADCP (akoestisch) en troebelheid (optisch) aan station Grens (boven) en Antwerpen (onder) voor meetcampagne 12-2015.
2.6.Monitoring Plankton en Fytobenthos
2.6.1.Totaal chlorofyl a en andere pigmenten
Substalen voor chlorofyl a en andere pigmenten komen uit hetzelfde schepstaal dat voor waterkwaliteit werd genomen. Een zo groot mogelijk volume water wordt over een 25 mm diameter GF/F glasvezel filter afgefilterd met als doel een zo hoog mogelijke gevoeligheid en nauwkeurigheid te kunnen bereiken. Per station zullen twee replicate filters gemaakt worden. De filters worden in het veld ingevroren in vloeibare stikstof zodat afbraak van het chlorofyl a vermeden wordt. Na transport naar het labo worden de filters in het donker bij -80°C bewaard, eveneens om afbraak van het chlorofyl te vermijden.
De analyses zullen ongeveer tweemaandelijks in batches van een 30-tal analyses gebeuren. Chlorofyl a verzameld op de filters zal geëxtraheerd worden door ultrasonnicatie met een tip-sonnicator in 10 ml 90% aceton gevolgd door 2 uur incubatie bij 4°C. Het pigmentextract wordt gefiltreerd over een 0.2 µm filter en geïnjecteerd op een High Performance Liquid Chromatography (HPLC) (Agilent 1100 Series) systeem.
Totaal chlorofyl a in de extracten zal gemeten worden aan de hand van HPLC (‘High Performance Liquid Chromatography’). Deze methode ondervindt in tegenstelling tot andere methodes (zoals spectrofotometrie of fluorometrie) geen interferentie van afbraakproducten van chlorofyl a zoals faeofytine of faeoforbide of andere chlorofyls zoals chlorofyl b of c. Dit is belangrijk in het kader van het uit te voeren onderzoek gezien dergelijke interfererende pigmenten in grote hoeveelheden in het estuarium voorkomen. Voor het scheiden van de pigmenten wordt de methode Van Heukelem & Thomas (2001) gebruikt. Chlorofyl a zal worden geïdentificeerd op basis van het absorptiespectrum met behulp van een fotodiode array detector (DAD) en door vergelijking van de retentietijd met die van commerciële standaarden. De hoeveelheid chlorofyl a wordt daarna gekwantificeerd na kalibratie van de HPCL met commerciële chlorofyl a standaarden. Zowel spectrofotometrische als de gevoeligere fluorometrische detectie zullen worden toegepast.
De gebruikte methode is erg gevoelig en laat toe concentraties lager dan 0.1 µg/l nauwkeurig te meten (afhankelijk van het volume dat gefilterd wordt, gewoonlijk > 250 ml in het Schelde estuarium). Chlorofyl a (Chl a) wordt gebruikt als proxy voor de fytoplankton biomassa en kenmerkende pigmenten zoals fucoxanthin, chlorophyll b (Chl b), lutein, zeaxanthin, peridinin en alloxanthin worden gebruikt om de verschillende fytoplankton groepen te onderscheiden.
2.6.2.Fytoplankton
Monsters voor de bepaling van de fytoplankton samenstelling zullen maandelijks op alle stations ingezameld worden. De bemonstering zal simultaan gebeuren in samenspraak met de uitvoerders van de andere percelen. Een monster van 100 ml wordt gefixeerd met Lugol en in het donker bewaard.
De analyses van de stalen zullen binnen de 3 maand na inzameling uitgevoerd worden.
Naast de pigmentanalyse wordt de fytoplankton gemeenschap ook geanalyseerd met behulp van een omgedraaide microscoop. Een 5 tot 10 ml substaal wordt geconcentreerd in een sedimentatie kamer volgens de methode van Untermohl. De cellen in de sedimentatie kamer worden geteld met een Zeiss Axiovert omgedraaide microscoop op vergroting 200x en 400x. Identificatie wordt op een zo hoog mogelijk taxonomische niveau gedaan waarbij ook een licht microscoop gebruikt wordt voor de correcte identificatie. Een vast aantal van 400 units worden geteld (CEN, 2006) om de fytoplanktonsamenstelling te bepalen. Met behulp van gepubliceerde biovolume berekeningen kunnen de getelde cellen omgezet worden in biomassa.
De soorten worden ingedeeld in de groepen diatomeeën en niet-diatomeeën. Op basis van literatuurgegevens en uitgaande van hun verspreidingspatronen in het estuarium worden de verschillende soorten ook ingedeeld als brakwatersoorten of zoetwatersoorten. Voor deze indeling zal gebruik gemaakt worden van literatuurgegevens betreffende de verspreiding van fytoplankton in het Schelde-estuarium (bvb. Muylaert & Sabbe 1999, Muylaert et al. 2000) en nieuw verzamelde data. Vervolgens wordt de totale biomassa van de categoriën brakwater diatomeeën, ander brakwaterfytoplankton, zoetwaterdiatomeeën en ander zoetwaterfytoplankton berekend door de biomassa’s van alle soorten in deze categorieën op te tellen. De uiteindelijke resultaten van de analyses zullen worden uitgedrukt in µg C l-1. Het totale fytoplankton is de som van deze vier categorieën.
De nauwkeurigheid van de analyse zal nagegaan worden door de totale fytoplanktonbiomassa bepaald aan de hand van celtellingen en biovolume-metingen te relateren aan de concentratie totaal chlorofyl a; indien beide analyses goed uitgevoerd zijn zou een goede correlatie moeten worden waargenomen.
2.6.3.Microfytobenthos
Monsters voor microfytobenthos zullen van op het land worden ingezameld op vijf slikken langs het Schelde estuarium tussen de Nederlands-Belgische grens en Gent (Figuur 2-3). Van iedere getijde plaat wordt op 3-5 verschillende plaatsen stalen genomen op een transect loodrecht op de waterlijn en binnen de hoog- en laagwaterlijn. Bij iedere staalname worden drie replicaten genomen en gepoold om kleinschalige variatie te voorkomen. Tijdens de staalname wordt de bovenste 2 mm sediment in situ bevroren met behulp van vloeibare stikstof en bemonsterd met een contact core. De bevroren stalen worden vervolgens in vloeibare stikstof bewaard tijdens het veldwerk en daarna bewaard bij -80°C tot aan analyse.
De analyses zullen in batches van een 45-tal monsters uitgevoerd worden en dit binnen de 3 maand na inzameling. Voor analyse wordt het sediment eerst gevriesdroogd om overtollig water te vermijden. Vervolgens worden de pigmenten uit het equivalent van ongeveer 1 ml sediment geëxtraheerd in 5 ml 90% aceton door sonnicatie met een tip-sonnicator gevolgd door 2 uur incubatie bij 4°C. Het extract wordt gefilterd over een 0.2 µm filter om partikels uit het extract te verwijderen.
De analyse gebeurt aan de hand van HPLC volgens de zelfde methode als voor de bepaling van totaal chlorofyl a, behalve dat in dit geval ook andere pigmenten bepaald zullen worden. Naast chlorofyl a, dat een maat geeft voor de totale biomassa van het microfytobenthos, zullen ook concentraties aan accessorische pigmenten bepaald worden. Deze geven een idee over de bijdrage van verschillende taxonomische groepen tot het microfytobenthos. Zo zal de concentratie aan chlorofyl c en fucoxanthine bepaald worden als indicatoren voor diatomeeën en chlorofyl b en luteïne als indicatoren voor groenwieren. Andere pigmenten die eventueel in hoge concentraties voorkomen en die een mogelijke indicatie kunnen geven over de samenstelling van het microfytobenthos zullen ook bepaald worden.
De gevoeligheid van deze methode is lager dan 0.2 µg pigment per ml sediment.
2.6.4.Pelagische primaire productie in het estuarium
Staalname voor de monitoring van primaire productie tijdens de vaartochten in het estuarium (pelagische primaire productie in functie van diepte) wordt uitgevoerd door UAntwerpen (zie 2.1.1), analyses door NIOZ.
Incubaties van stalen worden uitgevoerd in een incubator in een lichtgradiënt variërend van 0 tot 800 µmol photons m-s s-1 zoals beschreven door (Kromkamp en Peene 1995). De stalen worden gedurende ongeveer 2 uur geïncubeerd in duplo bij in situ temperaturen na toediening van een geringe hoeveelheid radioactief NaH14CO3 (bicarbonaat), en van elk monster wordt via een geautomatiseerde potentiometrische titratie de totale hoeveelheid opgelost CO2 bepaald. Hierdoor kan de specifieke activiteit worden berekend. Tijdens de incubatie worden de stalen langzaam geroteerd (3 Hz) om bezinking in de incubatiebuizen te voorkomen. De lichtintensiteit in de buizen wordt bepaald met behulp van een gekalibreerde sferische PAR (400 nm - 700 nm) sensor.
Na incubatie worden de stalen gefilterd en de filters worden minimaal 1 uur geïncubeerd in een damp van zoutzuur om niet gebonden radioactief bicarbonaat te verwijderen. Vervolgens wordt de radioactiviteit van de filters na toediening van een telvloeistof gemeten m.b.v. een scintillatietellen, waarbij een correctie voor quenching plaatsvindt. De donkerwaarden worden van de lichtwaarden afgetrokken om te corrigeren voor mogelijke bijdrage van chemo-autotrofe bacteriën.
De gemeten radioactiviteit wordt vervolgens omgerekend naar de gefixeerde hoeveelheid CO2 (mg/L/uur), met gebruikmaking van de specifieke activiteit. Door de data nu te fitten als een functie van de lichtintensiteit wordt een fotosynthese–lichtcurve (P-E) verkregen, waarbij de data worden genormaliseerd per mg chlorofyl-a (chla). Uit de verkregen fit kunnen dan de belangrijke parameters PBmax (mg C (mg chla)-1 uur-1 , de maximale fotosynthesesnelheid, en α, een maat voor de fotosynthese efficiëntie (mg C (mg chla)-1 uur-1)(µmol photons m-s s-1)-1 ) worden verkregen. In eerste instantie zullen de data worden gefit m.b.v. (Eilers and Peeters 1988):
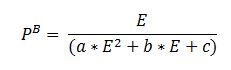
E is de lichtintensiteit en a, b en c zijn fitconstantes. Hierbij is PBmax 1/(2+√(ac) en α = 1/c.
De primaire productie is nu de integratie over de hoeveelheid chla per diepte-interval (aangeleverd via Perceel 1, basis waterkwaliteit), de lichtintensiteit en de fotische diepte, d.w.z. de diepte tot waarin nog fotosynthese mogelijk is, in dit onderzoek genomen als de 1% diepte van het licht aan het oppervlak. Als de fotische diepte groter is dan de werkelijke diepte wordt de laatste genomen. De lichtintensiteit E wordt berekend uit de diffuse lichtverzwakkingscoefficient kd (m-1):
![]()
Ez en Eo zijn de lichtintensiteiten op diepte z en aan het opprvlak respectievelijk. Hierbij zullen gemiddelde uurwaarden worden gebruikt voor Eo. De totale kolom productie is dus:
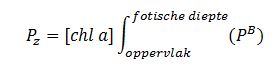
De dagelijkse primaire productie kan nu worden verkregen door de als boven berekende primaire uurproductie (mg C m-2 uur-1) te herhalen voor ieder uur van de dag en vervolgens deze uurproducties te sommeren tot een dagproductie (mg C m-2 dag-1).
2.6.5.Primaire productie microfytobenthos (benthische primaire productie)
Voor het microfytobenthos (bentische microalgen) wordt uitgegaan van 5 stations en 6 tijdstippen. Voor het microfytobenthos wordt de techniek gebruikt zoals beschreven in Barranguet & Kromkamp 2000; Barranguet et al. 1998; Kromkamp & Forster 2006. Hiertoe zullen een per station 3 random genomen oppervlakte stalen worden genomen. De bovenste mm van die stalen zullen worden gepooled tot 1 staal. Na verdunning met gefilterd stationswater zal de primaire productie worden bepaald op analoge wijze als de pelagische monsters, gebruik makend van het fotosynthetron (Figuur 2-16). De fotosynthese lichtcurves zullen op dezelfde wijze worden gefit als bij het fytoplankton.

Figuur 2-16 fotosynthetron
2.6.6.Zoöplankton
Tijdens elke campagne worden zoöplanktonstalen genomen, op volgende selectie van stations: Grens (Boei 87, km 57), Antwerpen, (km 78,5), Kruibeke (km 88), Temse (km 97,5), Lippenbroek (km 11), Dendermonde (km 121,5), Uitbergen (km 140), Melle (km 155), 3 stations in het Rupel bekken: Rupel (Boom), Beneden Nete en Dijle (estuarien) en 4 boundaries: Bovenschelde, Dender, Durme and Haven (Zandvlietsluis). Via schepstalen wordt 50 liter oppervlaktewater door een 50 µm net gefiltreerd. Het opgevangen zoöplankton wordt in een 200 ml plastiek pot gebracht, binnen de 12 uur verdoofd met spuitwater en uiteindelijk gefixeerd met formaline (4% eindconcentratie). Transport naar ‘Laboratoire Ecologie Fonctionnelle et Environnement’ (EcoLab; Université Paul Sabatier, Toulouse) gebeurt per verzendingsdienst of gebruik makend van verplaatsingen voor meetcampagnes.
Abundantie van de mesozooplankton organisme verschillende taxa wordt geteld onder binoculair microscoop (vergroting 90 x) en uitgedrukt in aantal individuen per m3. Microzoöplankton om de drie jaar geanalyseerd.
De grazing experimenten worden uitgevoerd aan de hand van incubatie-experimenten. Natuurlijk Scheldewater gefiltreerd op 250 µm wordt geïncubeerd met zooplankton (50 adulte E. affinis L-1) en zonder zoöplankton. De concentratie van de verschillende fytoplanktongroepen (essentieel diatomeeën en groenwieren) wordt gekwantificeerd na 24 u incubatie m.b.v. HPLC analyse, wat het kwantificeren van merkerpigmenten voor bepaalde algengroepen toelaat. De graasdruk op elke algengroep en de ingestie ervan worden berekend als clearance rate (Frost, 1972). Tegelijk met ieder incubatie experiment wordt ook een geconcentreerd zoöplankton staal bewaard in vloeibare stikstof. In het laboratorium worden E. affinis adulten uit deze stalen geïsoleerd geanalyseerd op darm pigmentinhoud, volgens de methode van Macka & Bohrer, 1076, aangepast voor het detecteren van selectiviteit.
Tendensen worden getest mat Spearmann – Rank, verschillen tussen series met Mann –Whitney, op een significantie niveau van minstens p< 0.05.
Een gedetailleerde analyse van de link tussen omgevingsfactoren en de zoöplankton spatio-temporele tendensen werd gemaakt voor de periode 2002-2012. Hierbij werd enkel gewerkt met de stations Melle (Z7), Uitbergen (Z7) en Dendermonde (Z6) om de invloed van saliniteit zo goed als uit te sluiten. Naast RDA analyses (CANOCO ) werd sterkte als predictor van de omgevingsfactoren op de verschillende zoöplankton taxa getest via GLM analyse met R.
Hoofdstuk 3.Klimatologisch overzicht
3.1.Inleiding
In dit overzicht worden de verklarende parameters temperatuur, verblijftijd en zout behandeld. Voor temperatuur werden de verschillende rekenparameters uit de Evaluatiemethodiek bepaald en besproken. Voor verblijftijd werd de evolutie van debiet beschouwd, voor zoutgehalte de trends in chloride.
3.2.Temperatuur
Over de ganse OMES periode valt geen significante stijging van de watertemperatuur op te tekenen: niet jaargemiddeld, niet per seizoen. Onderstaande figuur geeft een illustratie.
Figuur 3-1 Maandgemiddelde watertemperatuur voor de stations Grens en Dendermonde
Klimatologisch was 2015 een abnormaal warm jaar: de temperatuur te Ukkel was jaargemiddeld warmer dan normaal (www.meteo.be), de hoeveelheid zonneschijn was zeer abnormaal hoog. De seizoensgemiddelde watertemperatuur vertoont echter geen opvallende uitschieters in 2015. Enkel in de wintermaanden ligt de seizoensgemiddelde watertemperatuur in de Zeeschelde (op de zoete zone met korte verblijftijd na) hoger dan het gemiddelde over de voorbije omes periode (1996-2014), maar nog steeds onder het criterium uit de Evaluatiemethodiek (Tabel 3-1). In de andere seizoenen was 2015 koeler dan de voorgaande jaren. In de herfst overschrijdt de watertemperatuur in de gradiëntzone, net als de vorige jaren, wel het criterium van 15°C.
Tabel 3-1 Watertemperatuur in de verschillende zones van het estuarium. De in vet gedrukte temperaturen overschrijden het criterium uit de Evaluatiemethodiek

Wel opmerkelijk is de vervroeging van het moment waarop de temperatuur van 12°C wordt overschreden. Deze parameter werd in de Evaluatiemethodiek mee opgenomen als rekenparameter bij temperatuur: het moment waarop 12°C wordt overschreden kan indicatief zijn voor de start van het voortplantingsseizoen. Echter, sommige soorten worden eerder getriggerd door het lengen van de dagen, minder door toename van temperatuur. Zo kan een mismatch ontstaan tussen prooi en predator.
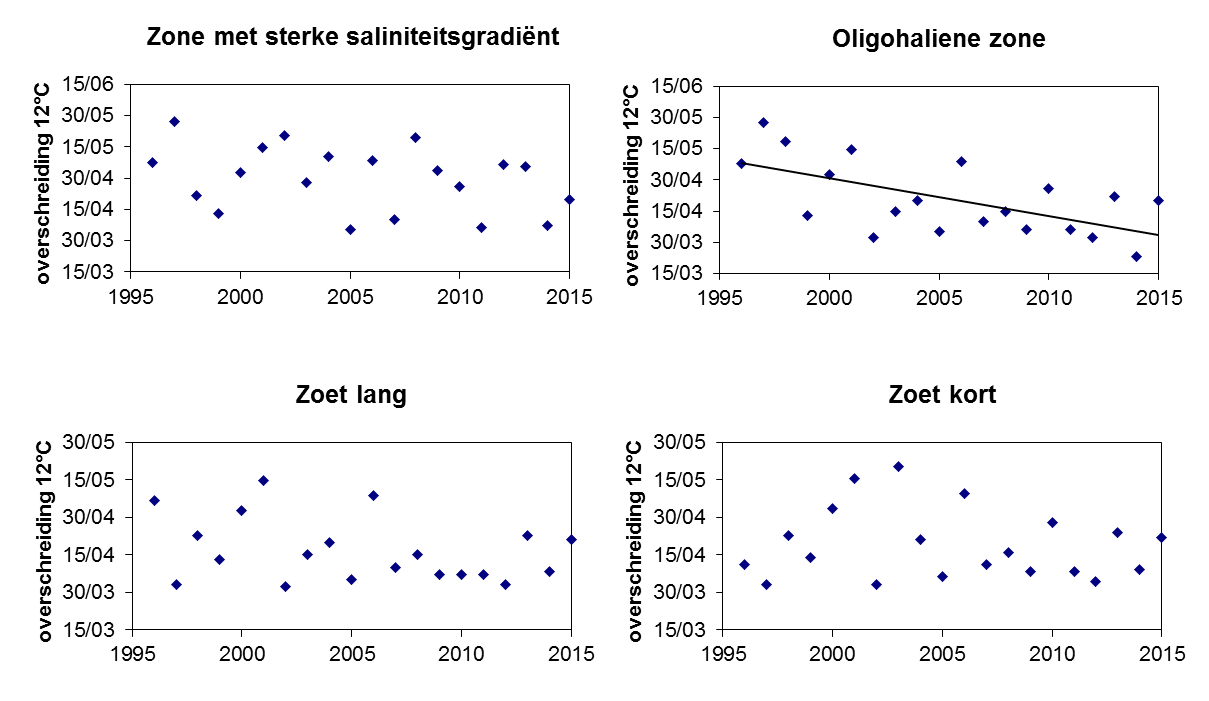
Figuur 3-2 Jaarlijks tijdstip waarop de eerste overschrijding van een watertemperatuur van 12°C werd geregistreerd in de verschillende zones van de Zeeschelde.
In de oligohaliene zone is er een significante (p < 0.01) vervroeging van het moment waarop de watertemperatuur 12°C overschrijdt. In de zone met sterke saliniteitsgradiënt en de zoete zone met lange verblijftijd is deze trend niet significant. In de zoete zone met korte verblijftijd is er geen trend.
Uiteraard wordt deze parameter beïnvloed door het moment waarop de vaarcampagne staat ingepland. Echter, de vervroeging in de oligohaliene zone is een maand, zodat dit niet volledig te verklaren valt door de inplanning van de staalname. Continue metingen van temperatuur kunnen uitsluitsel bieden: analyse van continue data voor het T2009 rapport toonde ook een duidelijke vervroeging van de overschreiding van 12°C. Of de trend zich ook in 2015 voorzet bij de continue temperatuursdata is nog niet gekend. Bij het opstellen van deze rapportage waren nog geen gevalideerde continue data voor 2015 beschikbaar.
Voorlopig is niet duidelijk of er effecten zijn op de planktonpopulaties.
3.3.Debiet en neerslag
Verblijftijd is een belangrijke verklarende parameter. Deze is in de Zeeschelde in hoofdzaak afhankelijk van het debiet. 2015 was een jaar met vrij normale winter en voorjaarsdebieten, maar met zeer lage zomer en najaarsafvoer (Figuur 3-3). Neerslaggegevens en de normalen voor neerslag werden overgenomen van het KMI (www.meteo.be). De normalen werden door KMI bepaald als het gemiddelde over de periode 1981 – 2010. Debietsgegevens werden verkregen van het HIC (www.waterstanden.be). Naar analogie met de normalen voor neerslag, werden de normalen voor debiet bepaald als het gemiddelde over de periode 1981-2010.
Figuur 3-3 Seizoensgemiddelde debieten te Schelle en Melle voor de periode 1996-2015, en de normale debieten te Schelle (data HIC) en neerslaggegevens en normale neerslag te Ukkel (data KMI).
Trends in winter- en lentedebieten lopen steeds mooi parallel met de hoeveelheid neerslag. In de zomermaanden is dit verband minder duidelijk. Wellicht speelt sluisbeheer hier ook een rol.
3.4.Chloride
Als maat voor het zoutgehalte worden binnen OMES de specifieke geleidbaarheid en het chloride gehalte gemeten. Op basis van het chloride gehalte werd het Schelde-estuarium ingedeeld in zones in de Evaluatiemethodiek (Figuur 2-4). De juiste ligging van deze grenzen is afhankelijk van getij en debiet (Figuur 3-4). In de zeer natte periode 2000-2002 is de zoutgrens ver stroomafwaarts verschoven. In de drogere zomer van bijvoorbeeld 2003 of 2011, schuift het zout verder opwaarts, met in het zoete af en toe concentraties die de eigenlijk de oligohaliene zone typeren.
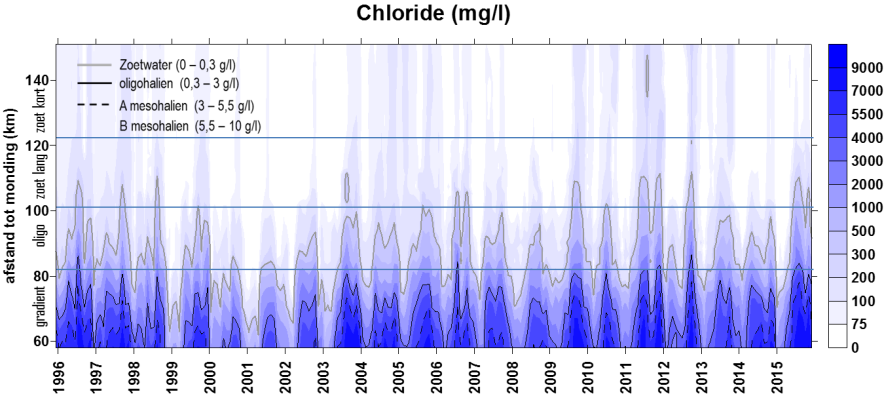
Figuur 3-4 Surface plot voor chloride, met aanduiding van de vaste Scheldezones (Gradiënt: zone met sterke saliniteitsgradiënt; oligo: oligohaliene zone; zoet lang: zoete zone met lange verblijftijd; zoet kort: zoete zone met korte verblijftijd). De contouren op de plot tonen de indeling in zoutklassen volgens het Venice systeem.
2015 was ook een jaar met een droge zomer en met perioden met uitzonderlijk lage debieten. De zoete zones zijn in 2015 dan ook iets minder zoet. In vergelijking met de zomer van 2009 – 2013, met gelijkaardig lage debieten, rukte het zout wel iets minder op.
Sinds 2009 worden bij het station Dendermonde (121 km van de monding) pieken in de chlorideconcentratie waargenomen (Figuur 3-5). De jaarlijkse piekconcentraties in 2009, 2010, 2011 en 2012 zijn 1.5 tot 2 keer hoger dan de maximale chlorideconcentraties die in tussen 1996 en 2008 werden waargenomen. Meer stroomafwaarts in de oligohaliene zone, bij station Temse (98.5 km van de monding), zijn dergelijke piekconcentraties niet uitzonderlijk: ze zijn het gevolg van een diepere zoutindringing in het estuarium bij lage bovenafvoeren. Echter, tot 2009 drong het zout bij lage bovendebieten niet verder door tot in Dendermonde. Sindsdien is dit wel het geval. Figuur 3-6 toont de correlatie tussen chloride en debiet, waarbij duidelijk vanaf 2009 veel hogere chloride gehalte kunnen voorkomen dan in de periode 1996-2008 bij gelijkaardige debieten.
De chloride pieken zijn niet gerelateerd aan veranderingen in zoutconcentratie aan de bovengrens van het estuarium. Integendeel, tot 2009 waren de chlorideconcentraties in het station Dendermonde sterk gecorreleerd met de concentraties in de Bovenschelde (Figuur 3-7): het chloride gehalte in de Bovenschelde bepaalde de concentratie in Dendermonde. Sinds 2009 is dit niet meer het geval en worden bij Dendermonde concentraties waargenomen die tot 3 maal de concentratie bedragen die verwacht werd uit het 1996-2008 verband tussen Bovenschelde en Dendermonde. Omgekeerd was er van 1996-2008 geen correlatie tussen de chloridegehaltes van Temse en Dendermonde wanneer de concentraties bij Temse groter waren dan 5 mM. Sinds 2009 is deze correlatie er wel: de hoogste chloride waarden te Dendermonde worden nu meer bepaald door afwaartse concentraties dan opwaartse condities. De invloed van de zee (zout) dringt dus dieper door in het estuarium.
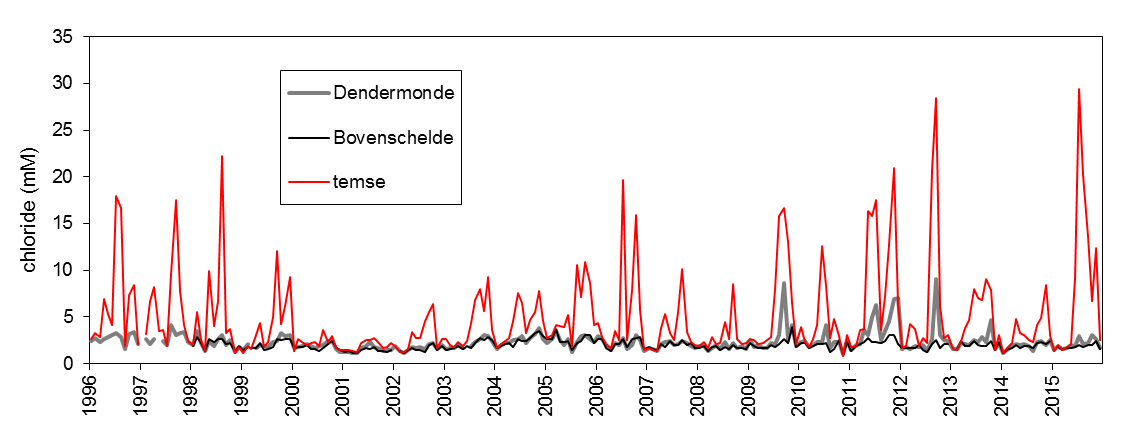
Figuur 3-5 Chlorideconcentraties opgemeten in de Boven-Zeeschelde (Dendermonde (km 121), Temse (km 98.5)) en de Bovenschelde.
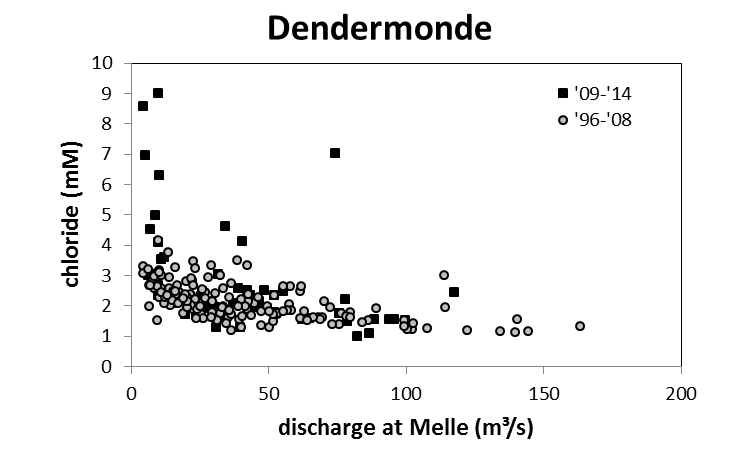
Figuur 3-6 Chloride gehalte (mM) te Dendermonde in functie van maandgemiddeld debiet (m³/s; data HIC)
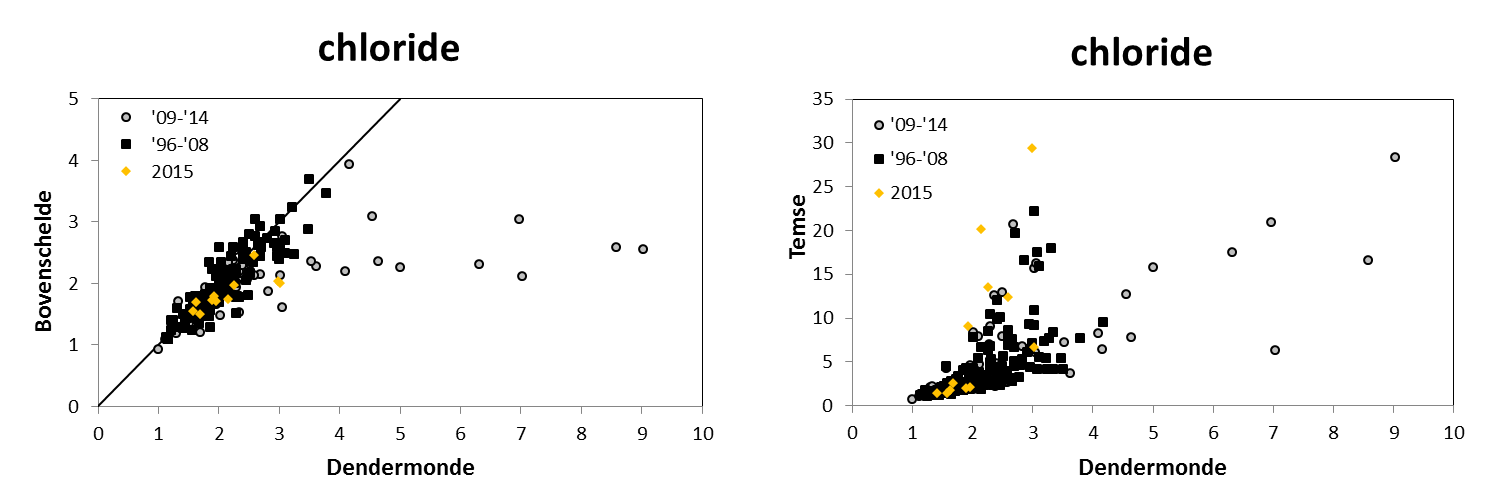
Figuur 3-7 Correlatie tussen de chlorideconcentraties opgemeten te Dendermonde en Bovenschelde (links) en de Temse (rechts).
Figuur 3-8 toont het verschil in chloride tussen Dendermonde en Bovenschelde. Voor 2009 was dit verschil minimaal en schommelde tussen ca +0.5 en -0.5 mM. Vanaf 2009 is dat verschil vele malen groter, en bijna steeds positief. Enkel 2014, een jaar met zeer hoge zomerdebieten, gedroeg zich “normaal”. 2015 kent terug positieve waarden die hoger liggen dan de periode voor 2009. Het verschil is echter klein, ondanks de zeer lage zomerdebieten.
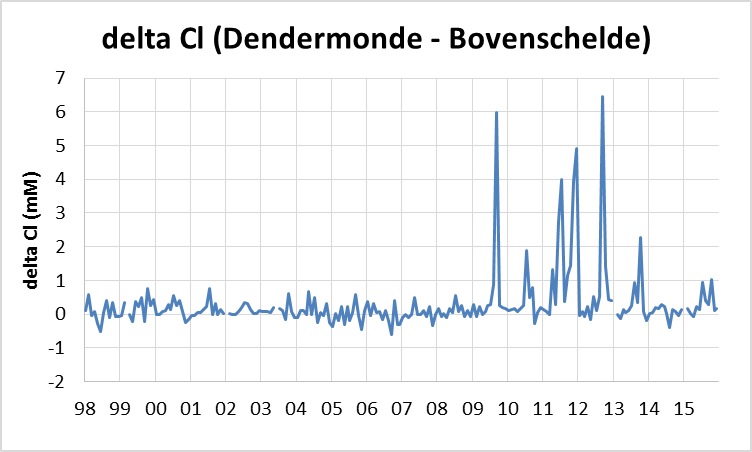
Figuur 3-8 Delta Chloride (verschil tussen chloride te Dendermonde en Bovenschelde) in de tijd.
Hoofdstuk 4.Basis waterkwaliteit
4.1.Zuurstof
4.1.1.Evaluatie 2015
Zuurstof is essentieel voor alle hoger leven, en dus een goede graadmeter voor de algemene waterkwaliteit van het estuarium. Zuurstof is daarom in de Evaluatiemethodiek opgenomen als toetsparameter voor waterkw. aliteit. Het is de resultante van zuurstofopname (door fysische beluchting en primaire productie door algen) en zuurstofverbruik (in hoofdzaak bacteriën). Sinds de start van de OMES metingen in 1995 zit zuurstof in duidelijk stijgende lijn (Figuur 4-1). Daar waar in de jaren ’90 alle zones in het estuarium te kampen hadden met ernstige zuurstoftekorten (< 2,5 mg/l), zijn deze problemen anno 2015 volledig van de baan (Tabel 4-1), op 1 tekort in het Rupelbekken na (2,36 mg/l in de Dijle op 5 mei). In 2006 wordt voor het eerst sinds de start van OMES oversaturatie gemeten, in de zoete Schelde. Anno 2015 komt oversaturatie hier frequent voor in de zomer. Vanaf 2010 komt ook in het Rupelbekken af en toe oversaturatie voor.
Tabel 4-1 Gemiddelde zuurstofwaarden per zone en toetsing aan de criteria uit de Evaluatiemethodiek. In zomer- en winterhalfjaar dient 95% (90% voor zone met sterke saliniteitsgradiënt) van de metingen boven 5 resp 6 mg/l te liggen. Alle metingen dienen boven 2,5 mg/l te liggen. In de gekleurde vakken worden de criteria niet gehaald.
|
Zone |
jaar | Zomer | Winter | ||||||
| gemiddelde | % norm | gemiddelde | % norm | ||||||
| O2 mg/l | % sat. | >5 mg/l | >2,5 mg/l | O2 mg/l | % sat. | >6 mg/l | >2,5 mg/l | ||
| gradiënt | 2015 | 7,59 | 80 | 100% | 100% | 9,01 | 77 | 100% | 100% |
| Oligohalien | 2015 | 6,39 | 68 | 89% | 100% | 7,96 | 68 | 100% | 100% |
| Zoet lang | 2015 | 6,94 | 73 | 97% | 100% | 8,99 | 76 | 95% | 100% |
| Zoet kort | 2015 | 7,87 | 82 | 100% | 100% | 9,90 | 84 | 100% | 100% |
| Rupelbekken | 2015 | 6,60 | 69 | 91% | 98% | 8,22 | 71 | 100% | 100% |
| Durme | 2015 | 6,66 | 71 | 82% | 100% | 8,91 | 73 | 86% | 100% |
Sterke oversaturatie duidt duidelijk op een overmatige algenbloei en het niet in evenwicht zijn van het ecosysteem. Deze zeer sterke algenbloei leidde tot op heden gelukkig niet tot typerende eutrofiëringsproblemen zoals nachtelijke anoxie of zuurstoftekorten wanneer de algenmassa afsterft. Na jaren van steeds verbeterende (stijgende) zuurstofconcentraties, is 2014 het eerste jaar met een daling van de gemiddelde zuurstofconcentraties, en dit in alle zones. 2015 scoort terug beter: enkel de oligohaliene zone, de Durme en het Rupelbekken (Figuur 4-2) voldoen niet aan de criteria uit de Evaluatiemethodiek (95% van de metingen > 5mg/l in het zomerhalfjaar).
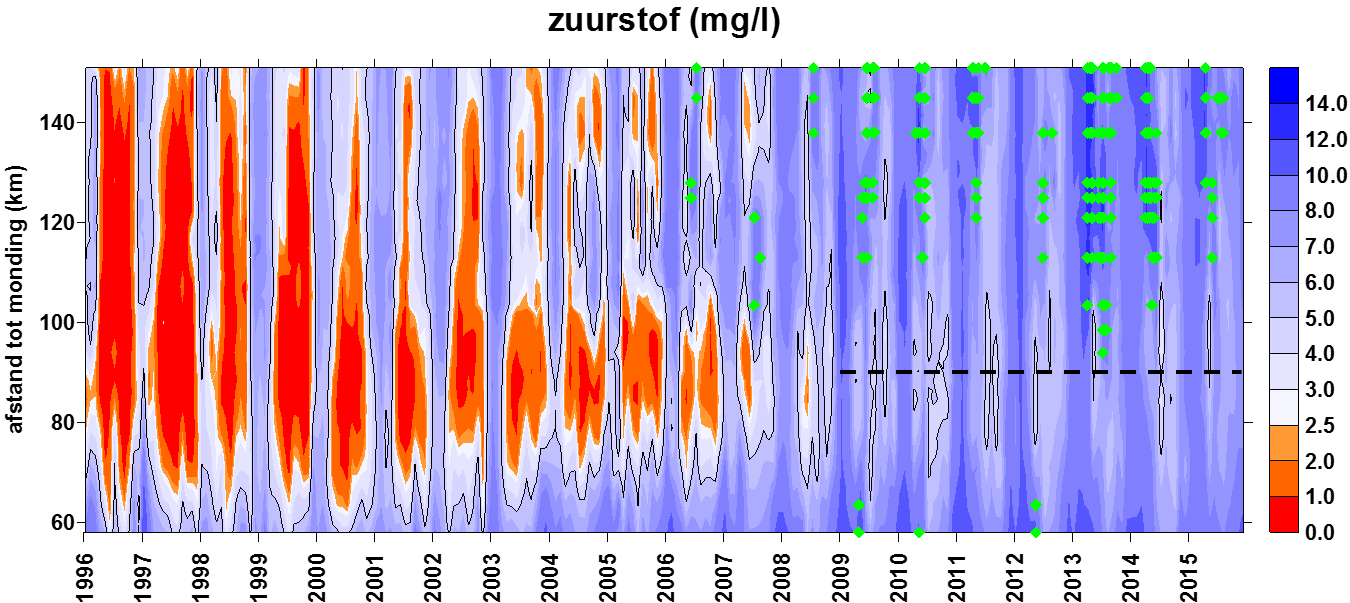
Figuur 4-1 Surface plot met zuurstofconcentratie (mg/l), op basis van maandelijkse metingen (winter) en twee metingen per maand (zomer vanaf 2009). De zwarte contouren markeren de grens van 5 mg/l. Rode en oranje waarden duiken onder 2,5 mg/l. Groene stippen zijn metingen met oversaturatie. De zwarte streeplijn duidt vanaf 2009 de Rupelmonding aan.
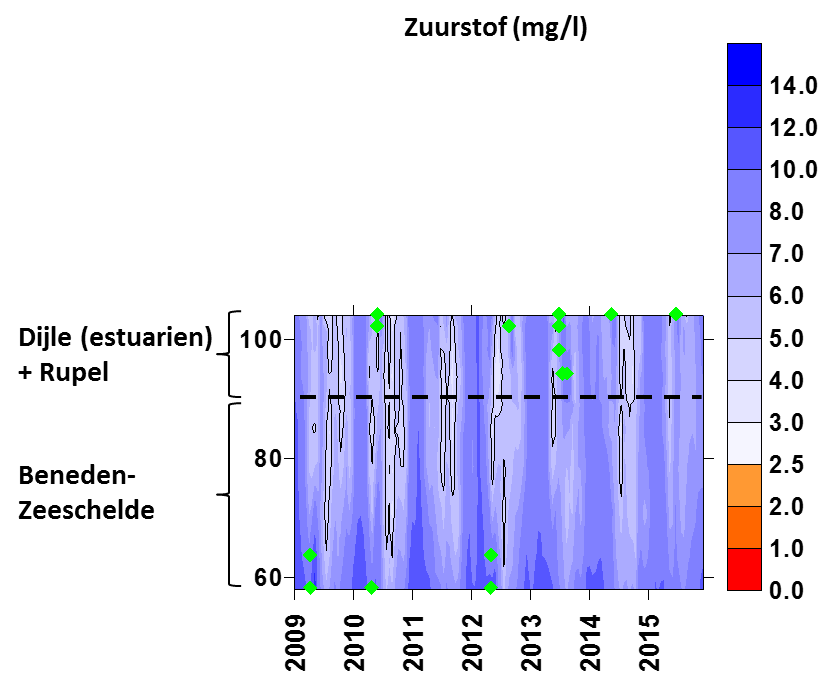
Figuur 4-2 Surface plot met zuurstofconcentratie (mg/l), op basis van maandelijkse metingen (winter) en twee metingen per maand (zomer) van Beneden-Zeeschelde via Rupel tot Dijle (estuarien). De zwarte contouren markeren de grens van 5 mg/l. Rode en oranje waarden duiken onder 2,5 mg/l. Groene stippen zijn metingen met oversaturatie. De zwarte streeplijn duidt de Rupelmonding aan.
4.1.2.Waterzuivering
Waterzuivering speelde een determinerende rol in de verbetering van het zuurstofgehalte en de algemene waterkwaliteit. Daarom werd de evolutie van de bouw van rioolwaterzuiveringsinstallaties (RWZI’s) voor het ganse Scheldebekken (Vlaanderen, Wallonië, Brussel, Frankrijk) opgelijst. De capaciteit van alle RWZI, uitgedrukt in inwoner equivalenten (IE), werd gesommeerd per Scheldezone om de invloed op de waterkwaliteit na te gaan (Figuur 4-3).
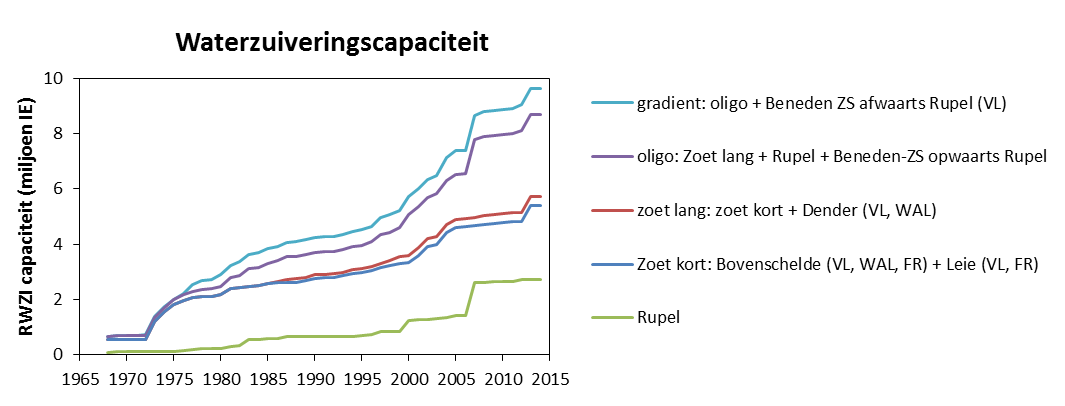
Figuur 4-3 RWZI capaciteit in het Scheldebekken, ingedeeld per Scheldezone. Bij elke Scheldezone staat aangegeven welke (deel)bekkens hierin uitmonden.
Uit Figuur 4-3 blijkt dat de RWZI capaciteit van quasi niets in de jaren ’70 toenam tot bijna 10 miljoen IE nu. Ook over laatste 20 jaar, de periode van de OMES metingen, was er ongeveer een verdubbeling van de capaciteit aan waterzuivering. De toename aan capaciteit vertaalt zich in een verbeterende waterkwaliteit. De toename aan capaciteit is echter niet steeds mooi lineair in de tijd en verloopt ook niet steeds even snel in elke zone. Daarom wordt het interessant om te kijken of de patronen in toenemende capaciteit ook terug te vinden zijn in de verbeterende waterkwaliteit.
Vanaf 1996 worden de OMES data, gemiddeld per zone, vergeleken met de RWZI capaciteit per zone. De effecten van vervuiling (en dus ook van waterzuivering) zijn meestal het sterkst voelbaar in de zomer, wanneer biologische processen actief zijn die onder andere het zuurstofgehalte naar beneden kunnen halen. Daarom worden hier de trends in zomerhalfjaargemiddelden besproken.
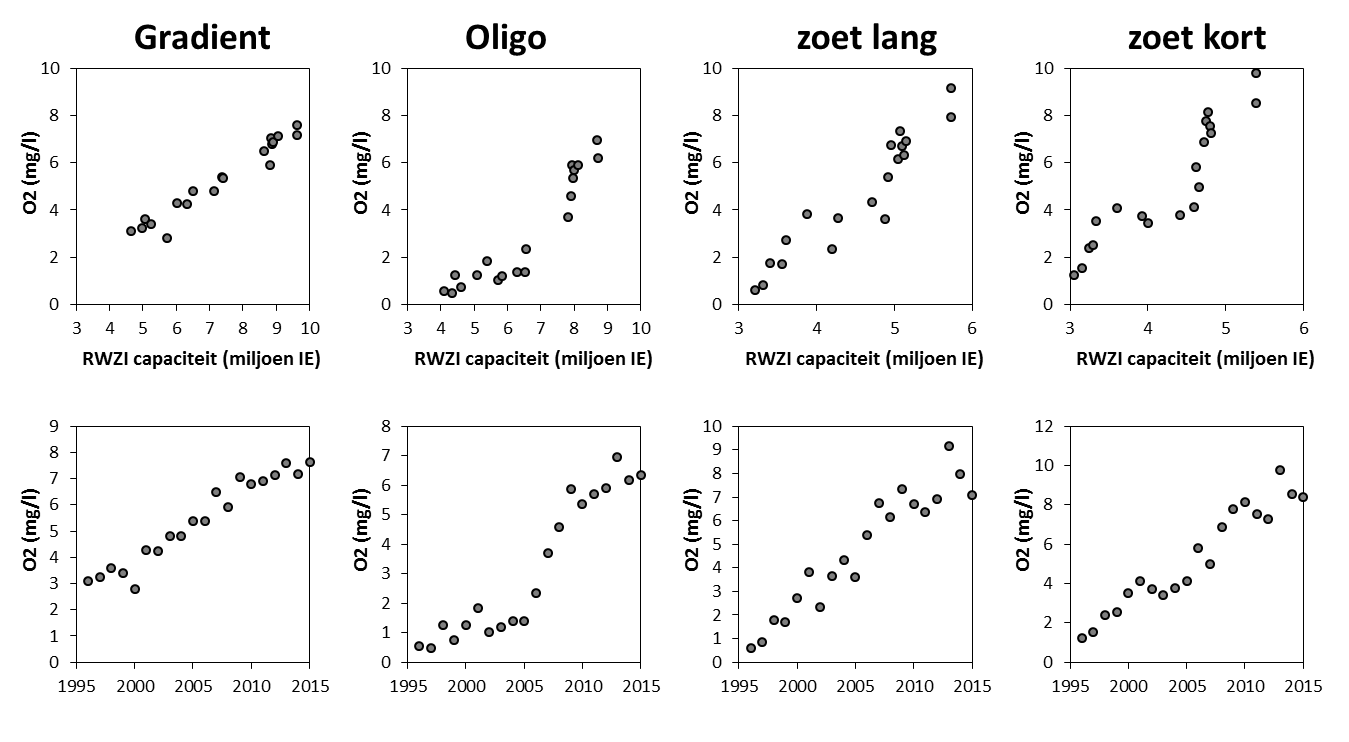
Figuur 4-4 gemiddelden per zone voor zomerhalfjaar
Zoete korte verblijftijd – zoete zone lange verblijftijd
De jaren ’90 worden gekenmerkt door zomers met quasi geen zuurstof in de zoete zones. Oorzaak is de veel te hoge vuilvracht komende vanuit de Bovenschelde (zoete zone korte verblijftijd), aangevuld met Dender (zoete zone lange verblijftijd), met zowel hoge CBOD als NOD (zie 4.1.3). Bovendien was het instromende water vanuit de bovenvermelde zijrivieren reeds arm aan zuurstof. Het zuurstofgehalte in de winter lag een stuk hoger: door hogere winterdebieten werd de vuilvracht enerzijds sterk verdund, maar vooral de lagere temperaturen maakten dat de bacteriële zuurstofvraag veel lager lag.
In de zoete zone met korte verblijftijd leverde een toenemende RWZI capaciteit aanvankelijk een sterke stijging van zuurstof op, om vervolgens niet meer toe te nemen bij toenemende RWZI capaciteit. Nochtans nam de inkomende vuilvacht, vanuit de Bovenschelde, wel quasi lineair af. Zowel NOD als BCOD daalden sterk (Figuur 4-5), ook in de periode waarin de verbetering van zuurstof in de Zeeschelde stagneerde. Vanaf 2003 stijgt zuurstof weer.
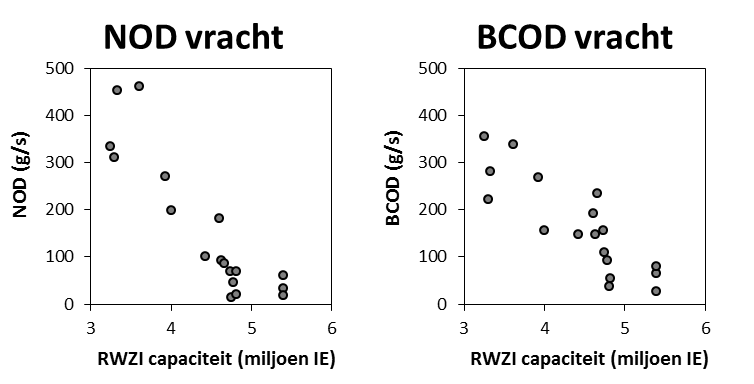
Figuur 4-5 jaargemiddelde NOD en BCOD vracht (g/s) in functie van RWZI capaciteit (miljoen IE) aan het boundary station Bovenschelde
De zoete zone met lange verblijftijd vertoont eenzelfde patroon. Bij zuurstof in functie van RWZI capaciteit zien we een verbetering, vervolgens stagnering om daarna sterk te stijgen.
De periode van stagnerende verbetering in de zoete Zeeschelde, gevolgd door een sterke stijging, kunnen we toeschrijven aan de shift van een stabiel zuurstofarm systeem naar een stabiel zuurstofrijk systeem, de zogenaamde regimeshift. In de jaren ’90 hield de grote vuilvracht de zoete Zeeschelde als het ware in een anoxische wurggreep: de vuilvracht onttrok zeer veel zuurstof uit het water. Lage zuurstofwaarden en hoge ammoniumconcentraties maakten dat zelfs algenbloei er onder te leiden had en er slechts een beperkte primaire productie en dus weinig extra zuurstofaanvoer mogelijk was. Voor 2003 was de zoete zone in een stabiele, heterotrofe toestand, waarbij respiratie de bovenhand haalt op zuurstofproductie en beluchting. Er was een aanzienlijke daling van de vuilvracht nodig om vanuit deze stabiele toestand in een andere stabiele toestand te komen: een autotrofe, zuurstofrijke toestand.
Het jaar 2003 is een keerpunt: de warme zomer van 2003 kent hoge temperaturen en lage debieten, en toch liggen de zuurstofwaarden hoger dan in de voorgaande zomers. Door de langzaam verbeterende waterkwaliteit in het bekken, werd in het estuarium een omslag bereikt. Sterke algenbloei werd mogelijk met meer zuurstofproductie tot gevolg. Meer zuurstof betekende snellere afbraak van de vuilvracht in het estuarium en betere condities voor algenbloei. Deze positieve feedback zorgde voor snelle stijgingen in het zuurstofgehalte. Op enkele jaren tijd verviervoudigden de zomerse zuurstofwaarden. De stijging ging wel met horten en stoten: in de zomer vertonen de zuurstofwaarden veel flickering: hoge en lage waarden wisselen elkaar snel af in de periode 2003-2007 (Figuur 4-6). Het ecosysteem switcht tussen twee alternatieve stabiele toestanden: een zuurrijke en een zuurstofarme. Door deze flickering is in de tijdreeks met zomergemiddelde zuurstofwaarden (Figuur 4-4) de sprong van een zuurstofarm naar een zuurstofrijk systeem niet opvallend aanwezig.
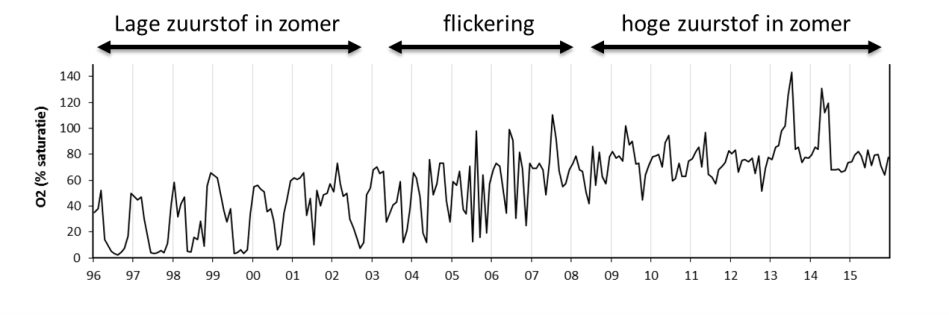
Figuur 4-6 Maandgemiddelde zuurstofverzadiging te Dendermonde
Vanaf 2008 kenmerkt de zoete Schelde zich door stabielere hoge zuurstofwaarden. Dankzij de dalende vuilvracht en de nog steeds zeer hoge primaire productie, stijgen de zuurstofwaarden verder. Vanaf 2007 verdwenen in de zone met lange verblijftijd de zuurstofwaarden onder 2,5 mg/l en was deze zone de eerste om te herstellen van acute zuurstoftekorten. De hoogste chlorofyl a waarden werden in de zoete zone met korte verblijftijd waargenomen. Dankzij deze sterke primaire productie die hier plaatsgrijpt, wordt de binnenkomende vuilvracht sneller gemineraliseerd. Dankzij deze processen in de zoete zone met korte verblijftijd, krijgt de zone met lange verblijftijd veel minder vuilvracht te verwerken. Daardoor werden, ondanks de hogere chl a waarden in de zone met korte verblijftijd, toch gemiddeld hogere zuurstofwaarden opgemeten in de zone met lange verblijftijd dan in de zone met korte verblijftijd. Af en toe duikt zuurstof nog onder 5 mg/l in de zomer in de zone met korte verblijftijd, ten gevolge van de mineralisatie van de binnenkomende vuilvracht. In 2012, 2013 en 2015 liggen echter meer dan 95% van de metingen wel boven 5 mg/l, waardoor wordt voldaan aan het evaluatiecriterium.
Oligohaliene zone
De Oligohaliene zone was de zone met de grootste zuurstoftekorten. Tot 2008 kwamen hier concentraties onder 2,5 mg/l voor. Deze zone kreeg eind jaren ’90 nog een grote vuilvracht vanuit de zoete zones doorgeschoven, plus via de Rupel de vuilvracht vanuit Brussel. Na de systeemomslag in de zoete zones, daalde de BOD-export van hier naar de oligohaliene zone sterk en ontving de oligohaliene zone water met een veel hoger zuurstofgehalte. Hierdoor zien we de oligohaliene zone dan ook verbeteren, zij het wel beperkter. In deze zone kwam immers nog steeds de ongezuiverde vuilvracht vanuit Brussel toe. Toch is de zuurstofconcentratie tussen 2005 en 2007 meer dan verdubbeld. Hoewel de waterzuivering in Brussel in 2007 werd opgestart, waardoor de RWZI capaciteit in de oligohaliene zone een sprong maakte van meer dan 1 miljoen IE, is deze verbetering toch voor een groot deel toe te schrijven aan het verbeterde ecosysteem in de zoete zones. Wellicht fungeerde de RWZI te Brussel in haar eerste jaar ook nog niet optimaal, want het is pas vanaf 2008 dat de effecten goed meetbaar zijn in de Rupel. Na 2008, wanneer effectief minder vuilvracht vanuit de Rupel komt, zet de stijging van het zuurstofgehalte zich sterk voort en verdwenen de acute lage zuurstofwaarden. In 2015 blijven alle waarnemingen boven 2,5 mg/l, maar toch blijft deze zone, samen met de Rupel, de zone met de laagste zuurstofwaarden in het estuarium. In 2015 voldeed slechts 89% van de metingen in de zomer aan het criterium. Op de surface plot is duidelijk te zien dat de zone met zuurstoftekorten zich zowel op- als afwaarts van de Rupelmonding manifesteert. Ondanks de waterzuivering in Brussel, ontvangt de oligohaliene zone nog steeds te veel vuilvracht, vooral NOD. Dit vergt veel zuurstof, maar de primaire productie ligt hier laag. In de oligohaliene zone is er waarschijnlijk zoutstress en is het lichtklimaat vaak ook ontoereikend. Het is de zone met gemiddeld de laagste chl a waarden, de primaire productie is er laag en de mortaliteit hoog, met af en toe zuurstoftekorten tot gevolg.
Rupel
Het effect van waterzuivering is spectaculair te zien op de Rupel (Figuur 4-7). Voor 2008 waren de zomerhalfjaar gemiddelden voor zuurstof steeds laag. Ondanks een bijna verdubbeling van de RWZI capaciteit in deze periode, van 0.8 naar 1.4 miljoen IE, bleef zuurstof laag en bleef ammonium hoog: een stabiel anoxisch systeem. In 2007 trad de RWZI Brussel-Noord in werking, maar het is wachten tot 2008 voordat de waterkwaliteit in de Rupel daadwerkelijk verbetert. Het effect van de RWZI is dan spectaculair: zuurstof verdubbelt van 2 naar meer dan 4 mg/l in 2008. Nadien stijgt zuurstof duidelijk verder, van 4 mg/l in 2008 naar meer dan 6 mg/l in 2015. Nochtans is in deze periode de RWZI capaciteit maar amper toegenomen, wat wijst op een duidelijk herstel van het ecosysteem functioneren na 2008.
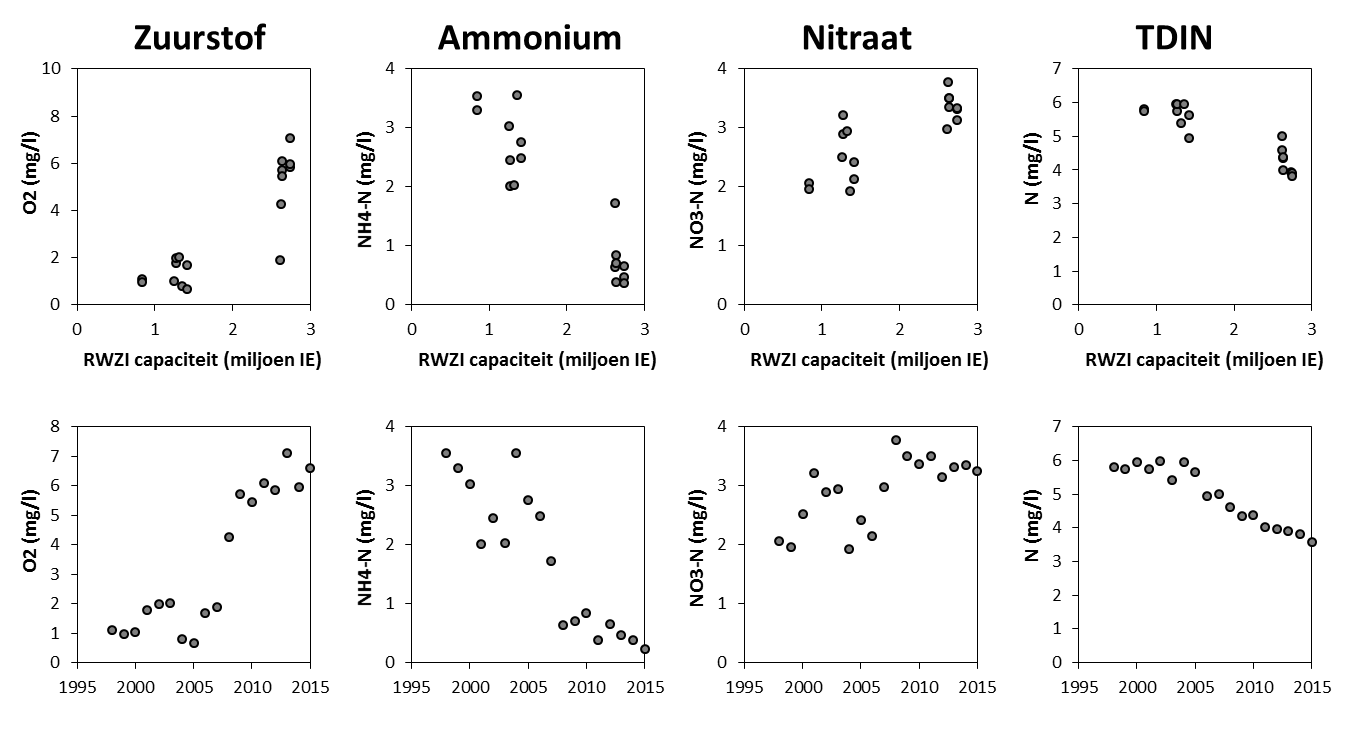
Figuur 4-7 gemiddelden voor de Rupel voor zomerhalfjaar
Zone met sterke saliniteitsgradiënt
De Zone met de Sterke Saliniteitsgradiënt was in het verleden nooit zo zwaar getroffen door lage zuurstofwaarden als de rest van de Zeeschelde. De grote verdunning met zuurstofrijker Westerschelde water lag aan de basis. Bovendien heeft het water reeds een lange weg afgelegd doorheen de opwaartse zones van de Zeeschelde, waar een groot deel van de vuilvracht werd verwerkt. Toch duurde het tot 2007 voordat acute zuurstofproblemen (zuurstof < 2,5 mg/l) verdwenen waren (Figuur 4-1). In 2015 wordt terug voldaan aan de voorwaarde voor een minimaal goed functioneren (volgens Evaluatiemethodiek): in de zomer ligt meer dan 90% van de metingen boven het zomercriterium van 5 mg/l, in het winterhalfjaar boven 6 mg/l. Merk op dat de evaluatiemethodiek in deze zone minder streng is dan in andere zones: daar dient 95% van de metingen te voldoen aan het criterium. Reden hiervoor is dat de Zone met Sterke Saliniteitsgradiënt van nature een zone is waar veel respiratie voorkomt, en dus lagere zuurstofwaarden niet ongewoon zijn. In de jaren ’90 was deze zone echter de zone met de hoogste zuurstofwaarden in de Schelde, niet omdat deze zone zo goed was, maar omdat de rest van de Zeeschelde er veel slechter aan toe was. Nu de Schelde terug aan de beterhand is, liggen de gemiddelde zuurstofconcentratie in de verschillende zones veel dichter bij elkaar.
In de zone met sterke saliniteitsgradiënt verloopt de stijging van het zomergemiddelde zuurstofgehalte zeer gelijkmatig, zowel in functie van de tijd als in functie van de RWZI capaciteit. Inzoomen op de stations Antwerpen en Grens toont echter wel een aantal interessante sprongen. Omdat voor deze stations langere tijdreeksen beschikbaar zijn, kan hier de evolutie op langere termijn geschetst worden. Voor het station Grens en Antwerpen werden voor de periode voor 1996 uit de OMES databank volgende datasets gebruikt: IHE, De Pauw, Afdeling Zeeschelde, RIKZ, VMM en CEME.
Antwerpen
Voor het station Antwerpen konden zomerhalfjaargemiddelden uitgezet worden vanaf de jaren ’60 (Figuur 4-8). Hoewel de bouw van de eerste 3 miljoen IE aan RWZI reeds zorgde voor een lichte verbetering, was bij het begin van de jaren ’80 de zuurstofconcentratie nog zeer slecht (< 2,5 mg/l). Tussen de jaren ’80 en het begin van de jaren 2000, de periode waarin de RWZI capaciteit verdubbelde van ca 3 naar 6 miljoen IE, verbeterde de zuurstofconcentratie in de zomer echter nauwelijks. De Zeeschelde kenmerkte zich als een heterotroof systeem met slechts beperkte zuurstofproductie en zeer sterke zuurstofconsumptie. De zuurstofvraag voor nitrificatie van de grote ammoniumvracht hield de zuurstofconcentratie laag. Parallel aan de toenemende RWZI capaciteit daalde de ammoniumconcentratie in deze periode echter zeer sterk (en steeg nitraat overeenkomstig). Vanaf ca 2003, wanneer ammonium een minimum bereikt heeft, stijgt het zuurstofgehalte spectaculair. In dat jaar start ook de omslag in de zoete Zeeschelde van een heterotroof naar een autrotroof systeem. Nitraat vertoont vanaf dan een daling.
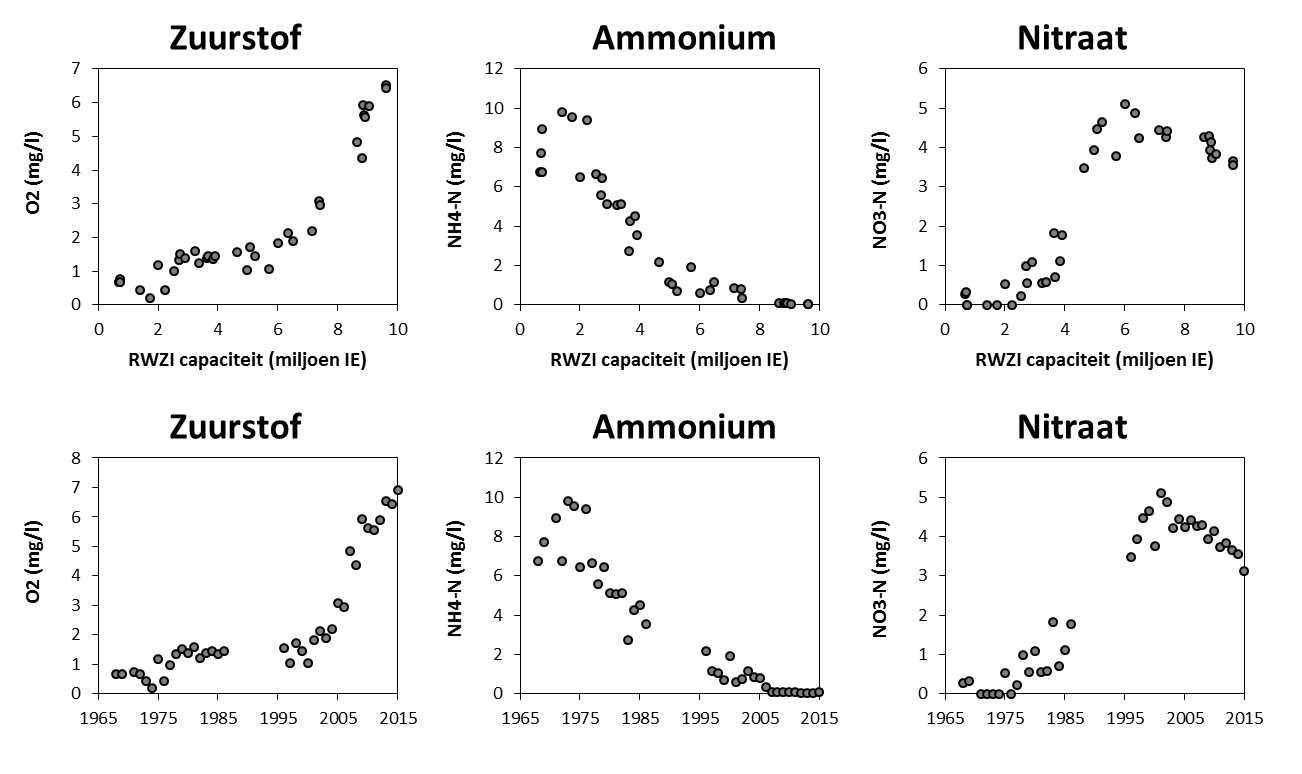
Figuur 4-8 Plots op basis van zomerhalfjaargemiddelden (april-september) aan station Antwerpen
Grens
De waterkwaliteit aan de grens is de resultante van alle processen in het estuarium en alle inspanningen in het bekken. Aan station Grens werden jaargemiddelde concentraties voor verschillende parameters uitgezet tegen de RWZI capaciteit enerzijds, en het jaartal anderzijds (Figuur 4-9). Zuurstof bereikt aan de Grens pas een triest minimum rond 1975. Tussen 1965 en 1975 neemt O2 af van ca 5 mg/l naar 1 mg/l; de bouw van RWZI’s in deze periode, tot 2 miljoen IE, kunnen de toenemende vuilvracht blijkbaar niet stoppen. Dit is ook de periode waarin de Antwerpse havenindustrie een grote expansie kent. Ammonium, de belangrijkste zuurstof vragende component in het water bereikt ook een maximum rond 1975. Vanaf dan zien we zuurstof spectaculair toenemen in de tijd, en met toenemende capaciteit aan RWZI’s. Toch vallen een paar sprongen op: de toename van 4 miljoen IE naar 6 miljoen IE gaat nauwelijks gepaard met een stijging van O2. Dit is de periode eind jaren ’90 – begin 2000, de periode waarin de zoete Zeeschelde nog door heterotrofie werd gedomineerd. Rond 2003 maakt zuurstof een plotse sprong van ca 6 naar ca 7 mg/l. Nochtans neemt in deze periode de RWZI capaciteit nauwelijks toe. Wat wel veranderde is de zoete Zeeschelde: zij schakelde over van een heterotroof naar een autrotroof ecosysteem, de zogenaamde regime-shift. In 2007 trad de waterzuivering te Brussel in werking, een verbetering met meer dan 1 miljoen IE. Aan station Grens levert dit echter geen sprong in zuurstof op.
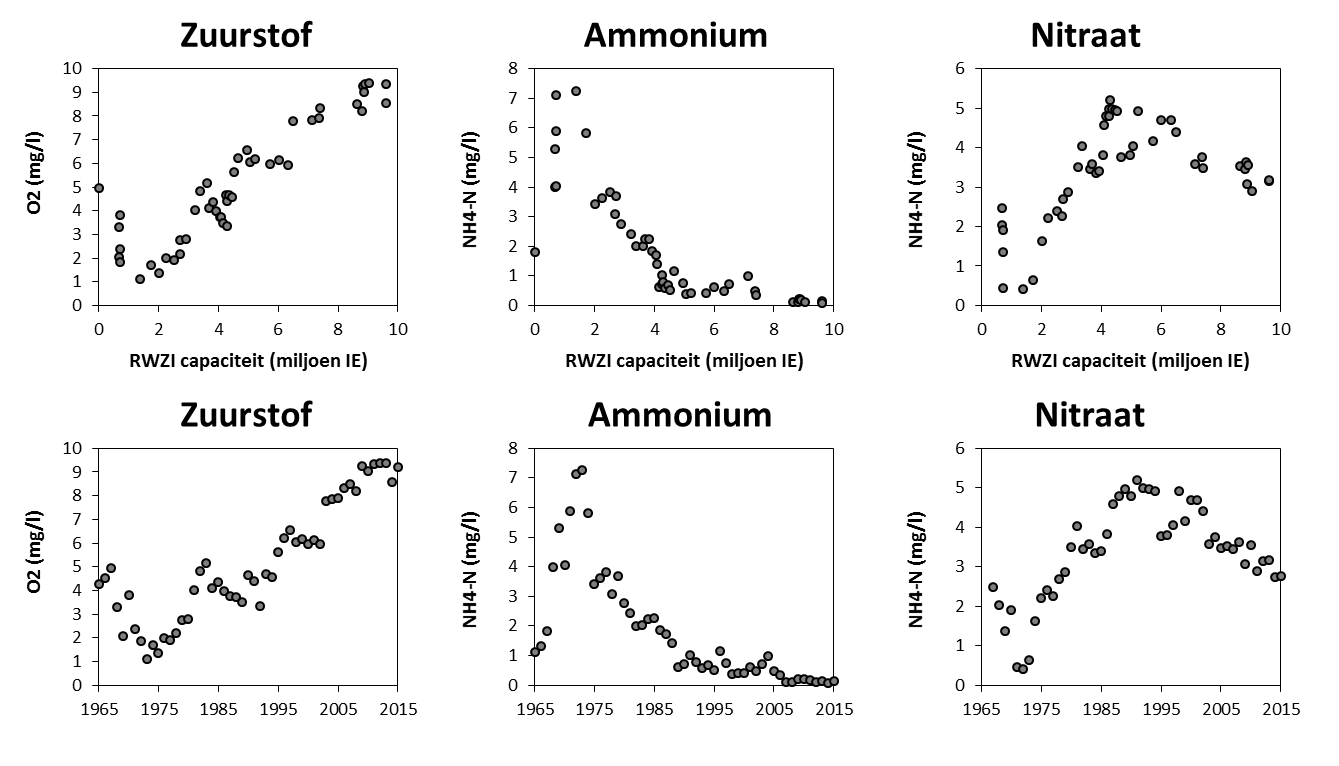
Figuur 4-9 Plots op basis van jaargemiddelden (april-september) aan station Grens
4.1.3.Zuurstofvraag: CBOD en NOD en koolstofdynamiek
Het zuurstofgehalte is de resultante van de zuurstofaanvoer enerzijds (primaire productie en fysische beluchting) en de consumptie anderzijds. De consumptie in de Schelde is hoofdzakelijk de respiratie door bacteriën, betrokken bij de verwerking van de organische afvalvracht en bij nitrificatie. De benodigde hoeveelheid zuurstof hiervoor wordt gemeten aan de hand van de biochemische zuurstofvraag (BOD). Het zuurstofverbruik in een BOD experiment is het resultaat van aerobe microbiële afbraak en mineralisatie van organisch materiaal (CBOD, carbonaceous biochemical oxygen demand, gelinkt aan de biodegradeerbare fractie van de organische koolstof) en van de nitrificatie van ammonium en nitriet (NOD, nitrogen oxygen demand) aanwezig in het waterstaal (zie ook hoofdstuk 2 Materialen en Methoden voor meer info).
Zoete zones
In de jaren ’90 bestond meer dan de helft van de totale zuurstofvraag uit zuurstofvraag voor nitrificatie (NOD) (Figuur 4-11). Vanaf 2000 zien we de NOD zeer sterk dalen, overal in het estuarium (Figuur 4-10). Deze daling is deels te danken aan de daling van ammonium in het bekken. De daling van NOD gaat echter sneller in het estuarium dan in de boundaries, en NOD neemt snel af in afwaartse richting. Dankzij de hogere zuurstofwaarden na 2000, kan een actieve populatie nitrificeerders het aanwezige NH4+ veel sneller omzetten dan in de jaren ’90, en zien we NOD langzaam verdwijnen uit het estuarium. Aanwijzing voor de aanwezigheid van een actieve populatie nitrificerende bacteriën vinden we in de correlatie tussen de theoretisch berekende NOD en de gemeten NOD (Figuur 4-12) (voor meer info, zie hoofdstuk 2 Materialen en Methoden). In het zomerhalfjaar, vanaf temperaturen boven 15°C, is nagenoeg alle NOD verdwenen uit het estuarium en voldoet de Zeeschelde aan het criterium van de Evaluatiemethodiek (2 mg/l). In de winter voldoen de zoete zones nog net niet: de aanvoer vanuit het bekken is nog te groot en de nitrificerende activiteit is in de winter te gering. Het is wellicht louter door verdunning dat de concentratie afneemt in afwaartse richting.
Ook de CBOD vertoont een dalende trend (Figuur 4-11), zij het iets minder spectaculair dan bij NOD. Toch is de CBOD concentratie in de zoete zone tussen 1996 en 2009 gehalveerd. De laatste jaren, wanneer de zoete Schelde vanaf 2009 in een stabiele, zuurstofrijke toestand is, is er van een verdere daling geen sprake meer maar vertoont CBOD eerder een zwak fluctuerend verloop (zie ook Figuur 4-13; vanaf 2010 zijn jaarrond CBOD data beschikbaar). Wellicht is de allochtone input van organische koolstof wel afgenomen, maar wordt de CBOD nu meer samengesteld door autochtoon geproduceerde koolstofverbindingen, ten gevolge van de sterke algenbloei. In de zomermaanden wordt hierdoor het criterium voor CBOD van 6 mg/l af en toe overschreden.
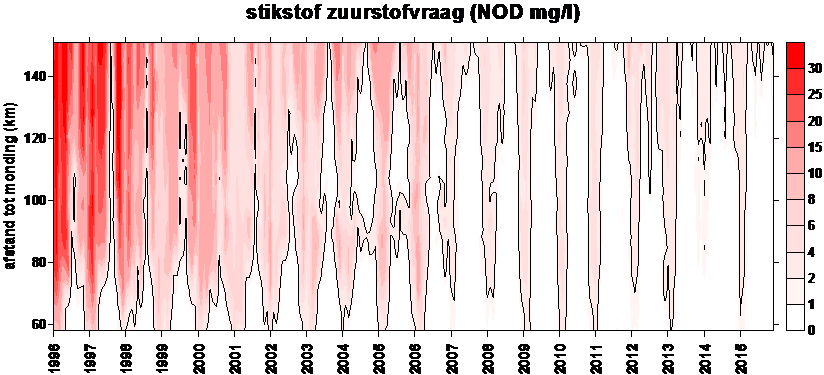
Figuur 4-10 Surface plot voor de biochemische zuurstofvraag voor nitrificatie (NOD mg/l), berekend op basis van NH4 en NO2. De zwarte lijn markeert de grens van 2 mg/l (criterium Evaluatiemethodiek)
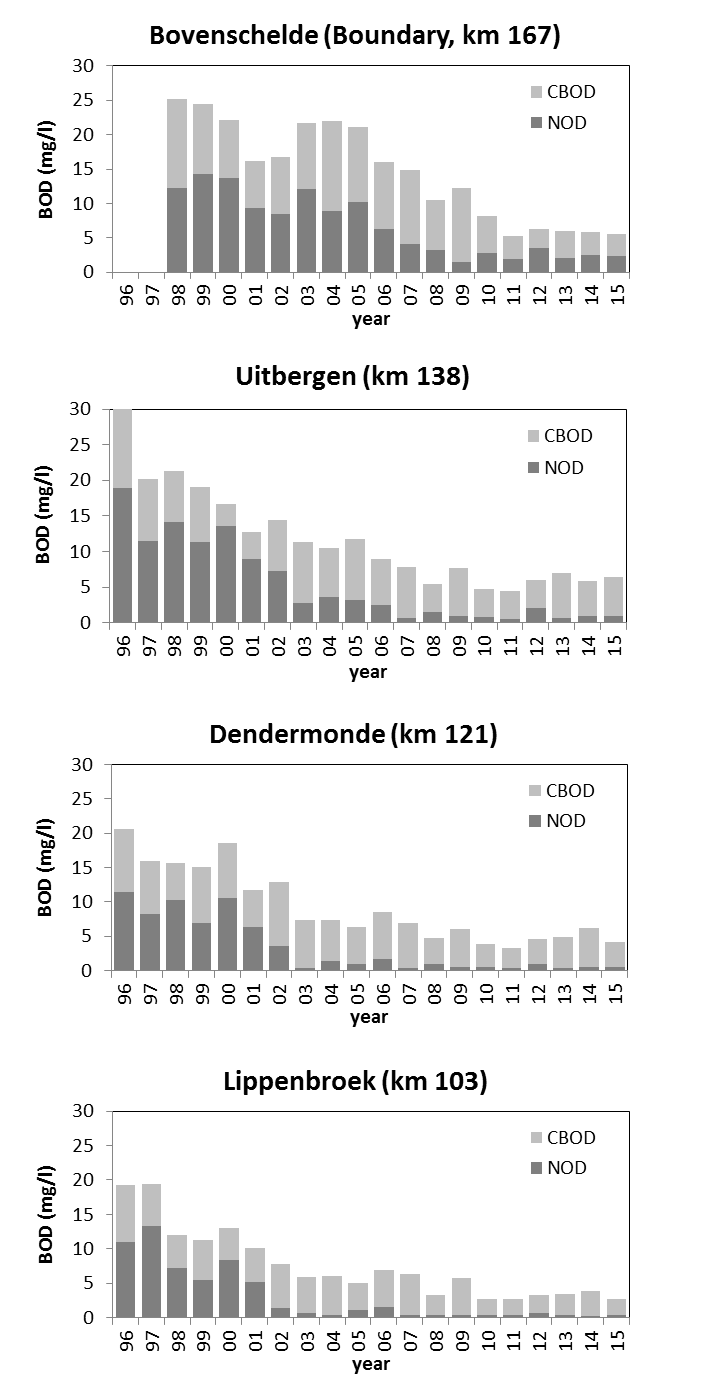
Figuur 4-11 Biochemische zuurstofvraag (BOD), opgesplitst in zuurstofvraag voor nitrificatie (NOD) en zuurstofvraag voor koolstofmineralisatie (CBOD), bij watertemperaturen boven 15°C.
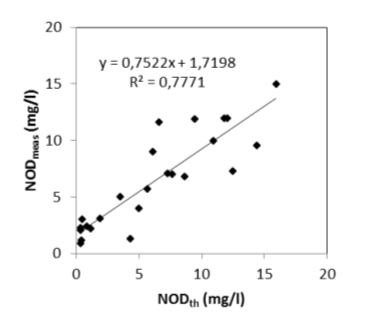
Figuur 4-12 Correlatie tussen NODth en NODmeas aan het station Dendermonde voor watertemperaturen boven 15°C.
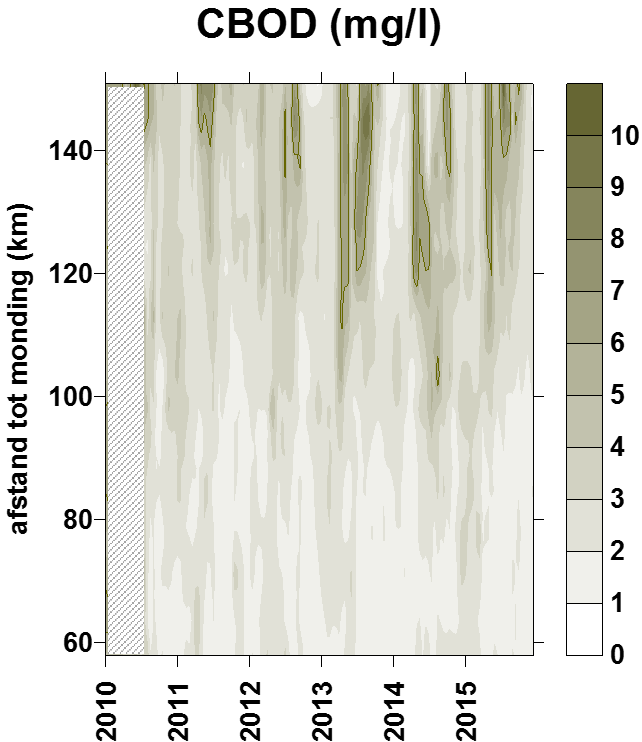
Figuur 4-13 Surface plot voor de biochemische zuurstofvraag voor koolstofmineralisatie (CBOD mg/l). De donkere lijn markeert de grens van 6 mg/l (criterium Evaluatiemethodiek).
Om inzicht te verwerven in de samenstelling van de CBOD, worden binnen OMES koolstofisotopen bestudeerd. De stijging van het gehalte autochtoon geproduceerde koolstofverbindingen wordt bevestigd door een stijgende 13C/12C isotopenverhouding in de opgeloste anorganische koolstof (δ13C-DIC) in het Scheldewater. Zoetwater fytoplankton (in tegenstelling tot marien fytoplankton) consumeert bij voorkeur CO2 met het lichtere isotoop 12C boven het zwaardere 13C. Hierdoor wordt bij sterke algenbloei in het zoete de opgeloste anorganische koolstof (DIC) in het Scheldewater armer aan 12C en stijgt bijgevolg de δ13C-DIC. Echter, een verminderde respiratie (door minder CBOD) kan hetzelfde effect hebben.
Wanneer als gevolg van sterke primaire productie de particulaire organische koolstof (POC) in de zoete Schelde veel algen bevat, zal deze POC rijker zijn aan het lichtere isotoop 12C, en daalt bijgevolg δ13C-POC. In theorie kunnen we de δ13C-POC van de fytoplankton biomassa geproduceerd in de Schelde schatten uit de δ13C-DIC metingen. De gemiddelde fractionatiefactor tussen substraat (DIC) en product (POC) voor koolstofopname door fytoplankton in de Schelde werd geschat op -22.3‰ (Hellings, 2000). Dat wil zeggen dat het organisch koolstof (POC) geproduceerd door fytoplankton door de opname van DIC met -22‰ lichter wordt dan het substraat DIC (omdat fytoplankton een voorkeur heeft voor het lichtere 12C). De δ13C-DIC waarden in de Schelde liggen tussen -14 en -11‰, de δ13C-POC van fytoplankton kan dus geschat worden op waarden tussen -36 tot -33‰. We zien echter niet echt een dalende trend voor de jaarlijkse gemiddelden van δ13C-POC (gemeten sinds 2006) wat er op wijst dat het particulaire organisch materiaal gemiddeld nog steeds grotendeels uit allochtoon materiaal bestaat. Dit allochtoon materiaal is aangerijkt in 13C (gemiddeld -27.4‰ volgens Hellings 2000) doordat het afkomstig is uit terrestrische planten (tussen -31 en -26 ‰ voor meest voorkomende vegetatie aan de Schelde), en afvalwater – gezuiverd of niet (tussen -26 en -28‰). Alleen de zomers en vooral de lente worden in het zoete gekenmerkt door een lagere δ13C-POC (Figuur 4-14).
Bovendien is er een stijgende trend van de ratio chl a/POC tussen de jaren 1990 en 2005 (Figuur 4-15) die ook suggereert dat de fractie autochtoon materiaal in de organische koolstof eerst gestegen is. Na 2005 daalt deze ratio dan weer.
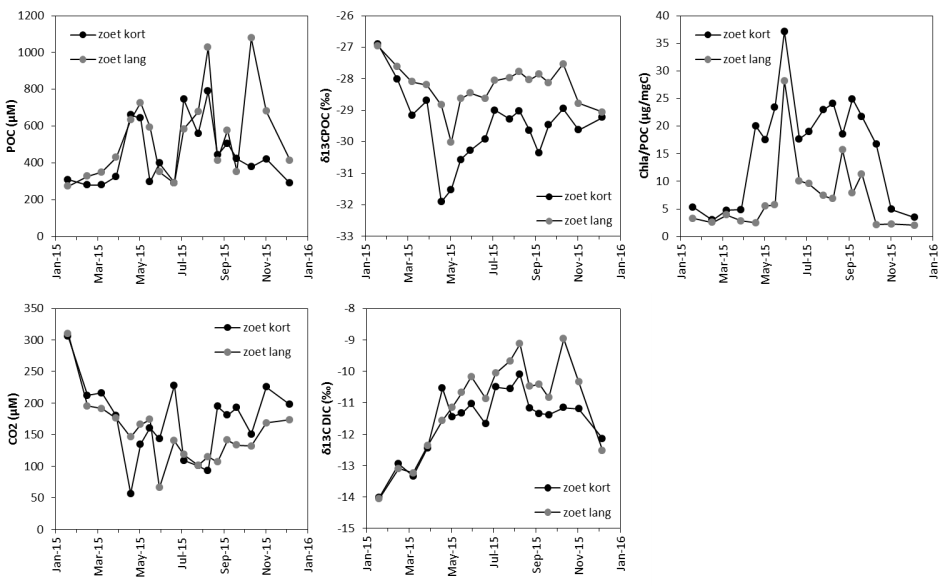
Figuur 4-14 Gemiddelde POC, δ13C-POC, Chla/POC, CO2, en δ13C-DIC in de zoete zones met korte en lange verblijftijden in 2015.
Door de toenemende zuiveringsgraad in het estuarium, daalde aanvankelijk dus de CBOD, een daling die wellicht door productie van autochtoon organisch materiaal stagneerde. De particulaire organische koolstof (POC) kent immers ongeveer eenzelfde verloop (Figuur 4-15).
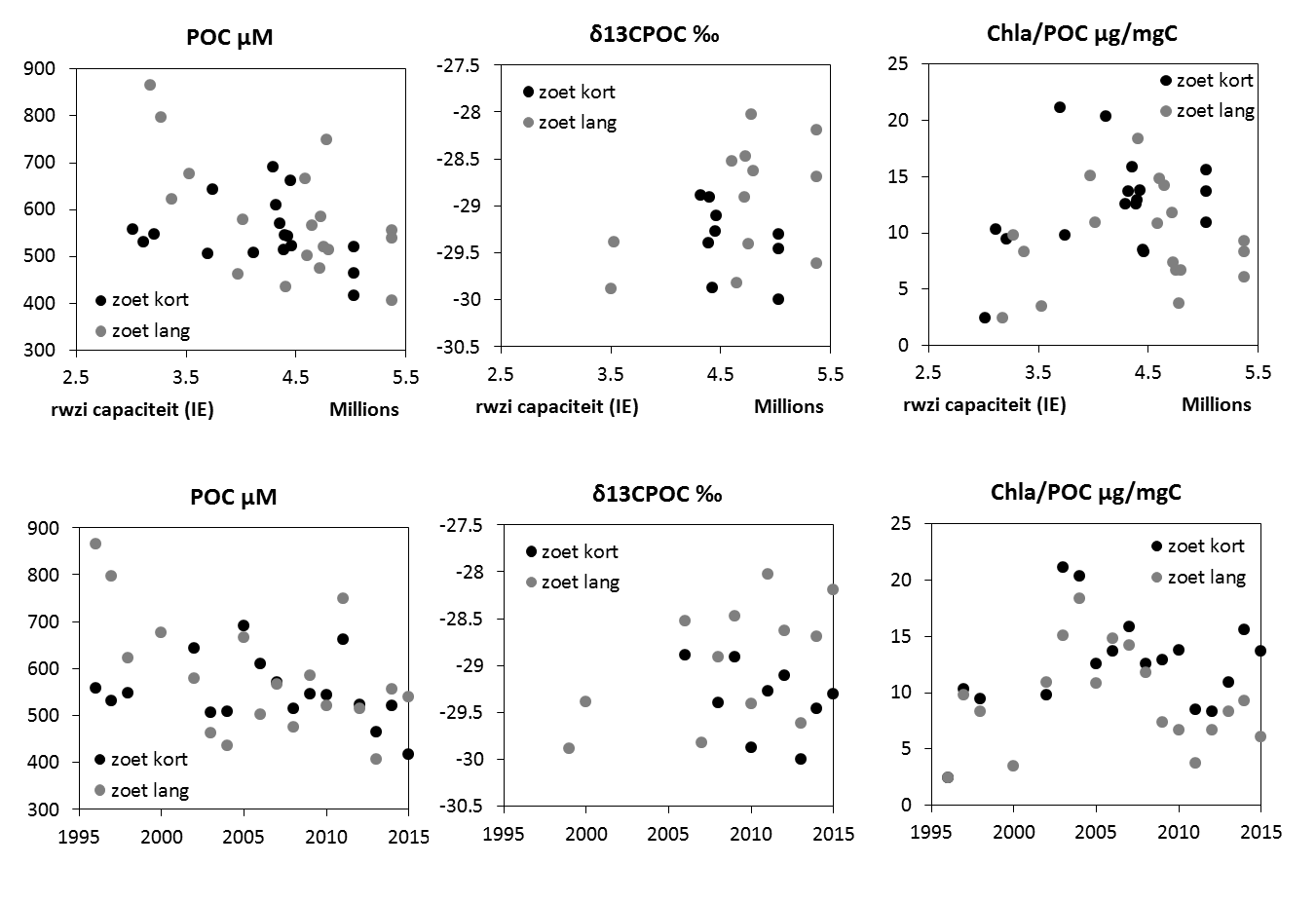
Figuur 4-15 Jaarlijkse gemiddelde POC, δ13C-POC en Chla/POC ratio in de zoete zone met korte en lange verblijftijden
In de zoete Zeeschelde stijgt δ13C-DIC duidelijk van het begin van de metingen tot op heden (Figuur 4-14). De CO2 consumptie door algen neemt duidelijk toe en er is ook minder respiratie (dalende CBOD), waardoor de δ13C-DIC omhoog gaat. Van Melle richting Grens wordt eenzelfde trend waargenomen: CO2 wordt opgenomen door algen en verdwijnt gaandeweg meer en meer uit het water. Het resterende CO2 wordt rijker aan het lichtere 12C waardoor δ13C-DIC stijgt.
Zoals eerder beschreven, consumeerde de verwerking van de grote CBOD vracht in de zoete Zeeschelde veel zuurstof. Organische koolstofverbindingen werden door bacteriën omgezet in CO2. Het estuarium kende dan ook een grote uitstoot aan CO2 in de zoete zone (Figuur 4-17). Met de halvering van de CBOD tussen 1996 en 2009, is ook de CO2-uitstoot gehalveerd. Vanaf 2009 vertoont deze, net zoals CBOD, geen dalende trend meer.
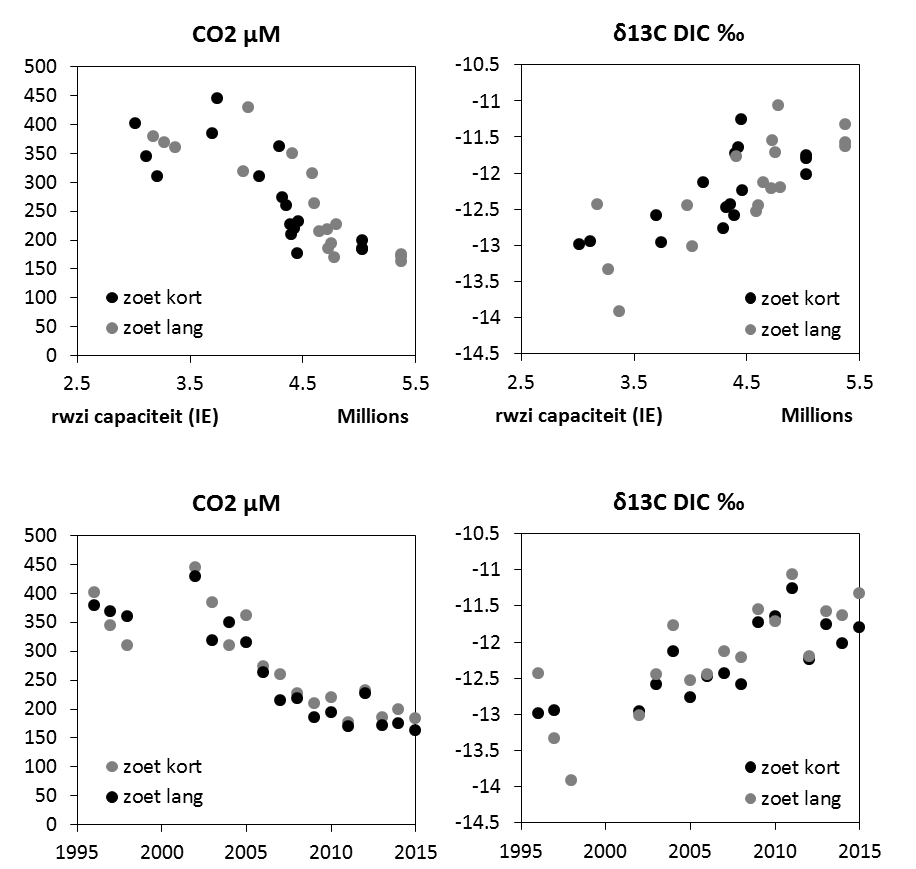
Figuur 4-16 Jaarlijkse gemiddelde δ13C-DIC en CO2 in de zoete zone met korte en lange verblijftijden
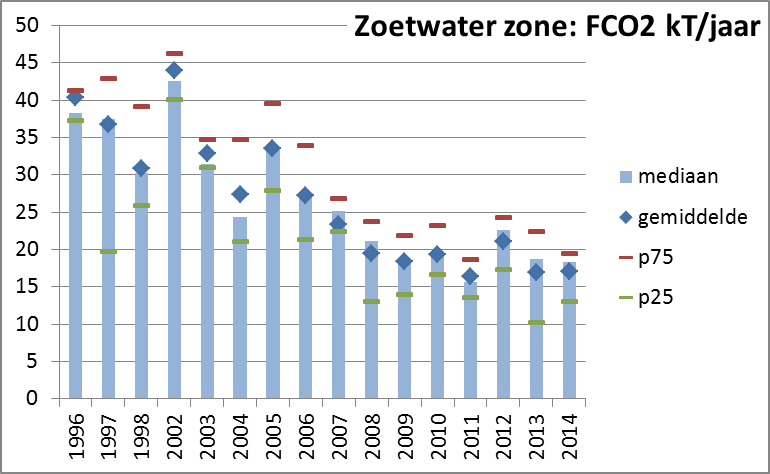
Figuur 4-17 CO2 uitstoot in de zoete zones van de Zeeschelde (kT/jaar)
Oligohaliene zone en zone met sterke saliniteitsgradiënt
In de oligohaliene zone en zeker in de zone met sterke saliniteitsgradiënt liggen de CBOD en NOD lager. Dit komt enerzijds doordat een deel van de vuilvracht in de zoete zones reeds werd verwerkt, anderzijds door toenemende verdunning met zeewater.
De concentraties aan CBOD liggen in deze zones wel lager, de totale vracht die hier passeert is net door het grote volume nog altijd hoog. De totale CO2-uitstoot in deze zones is dan ook hoog (Figuur 4-20). In de periode 1996-2003 lag de uitstoot van CO2 hier hoger dan in het zoete, door de grotere uitwisselingsoppervlakte, en de hoge CBOD en CO2 invoer uit het zoete. Door de slechte zuurstofcondities in het zoete stroomde immers meer CBOD door naar de oligohaliene en gradiëntzone. In de gradiëntzone was toen voldoende zuurstof aanwezig voor een sterke CBOD afbraak en sterke CO2-uitstoot. Na de systeemomslag werd meer CBOD verwerkt in het zoete, en daalde de CO2-uitstoot in de brakke zones sterk. De laatste jaren ligt de uitstoot in het zoete gemiddeld iets hoger dan in het brakke.
De lagere CBOD in de oligohaliene zone en in de zone met sterke saliniteitsgradiënt zien we ook terug in lagere POC concentraties. Naar de grens toe neemt POC af. Ook in de tijd neemt POC sterk af in de oligohaliene zone. In de gradiëntzone wordt geen daling gemeten. Wel verandert hier de isotoopsignatuur van de POC: δ13C-POC stijgt met ± 1.5‰. Dit kan een gevolg zijn van enerzijds de stijgende δ13C-DIC (±1‰ tussen 2005 en 2015) waardoor autochtoon geproduceerd POC ook met 1‰ aangerijkt wordt, en anderzijds een stijgende fractie van allochtoon materiaal.
De ratio ligt hier ook hoger dan in de zoete delen van het estuarium. Dat kan wellicht verklaard worden door een toenemend aandeel marien organisch materiaal (gemiddeld -22‰ in Vlissingen).
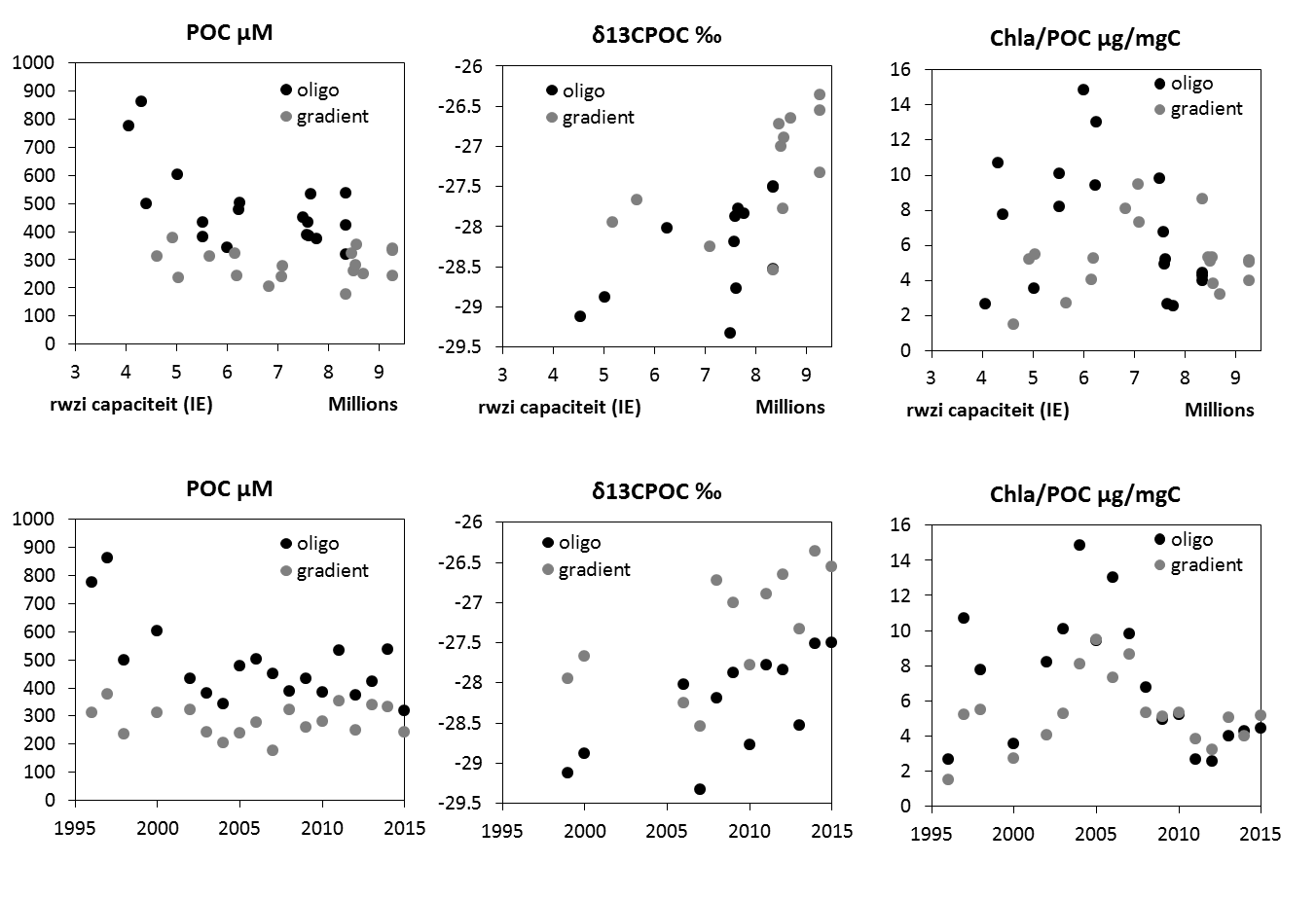
Figuur 4-18 Jaarlijkse gemiddelde POC, δ13C-POC en Chla/POC ratio in de oligohaliene zone en zone met sterke saliniteitsgradiënt.
In de oligohaliene zone en de zone met sterke saliniteitsgradiënt konden geen betrouwbare correlaties gevonden worden tussen NODmeas en NODth, wellicht omwille van lagere concentraties aan CBOD en NOD en dus grotere relatieve fout. Wel lagen de metingen vanaf 15 °C zonder nitrificatieremmer (BODwithout) meestal duidelijk hoger dan zonder nitrificatieremmer (CBOD), wat wijst op de aanwezigheid van een actieve populatie van nitrificerende bacteriën.
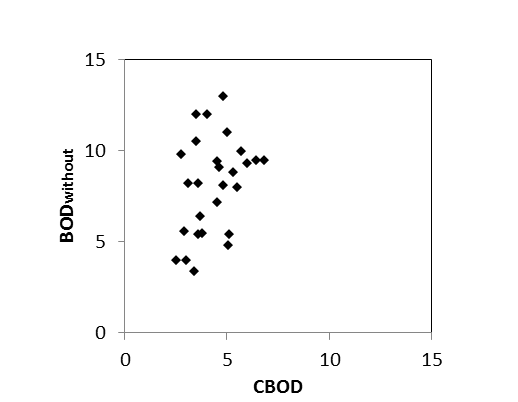
Figuur 4-19 . correlatie tussen CBOD en BODwithout in de oligohaliene zone (Kruibeke), bij temperaturen boven 15°C
Figuur 4-20 CO2 uitstoot in de oligohaliene zone en zone met sterke saliniteitsgradiënt (kT/jaar)
In de zomermaanden is in de oligohaliene en gradiëntzone quasi alle NOD verdwenen uit het water, en wordt ruimschoots voldaan aan criteria. In de winter daarentegen overstijgt NOD nog af en toe 2 mg/l. Door de lagere wintertemperaturen wordt de NOD niet of zeer traag verwerkt. Deze te hoge stikstof zuurstofvraag veroorzaakt in de winter dan ook eigenlijk geen problemen in de Zeeschelde.
Rupel
Probleemzone blijft nog steeds het Rupelbekken, en meer bepaald de vuilvracht die vanuit Zenne het estuarium binnenkomt. Die daalde spectaculair na de inwerkingtreding van de RWZI Brussel-Noord, een daling die zich nog langzaam verder zet (Figuur 4-21). Maar ondanks dat, blijven anno 2015 zowel CBOD als NOD zeer hoog. De NOD ligt jaargemiddeld ca 4 maal hoger aan de boundary Zenne dan aan Bovenschelde. Figuur 4-22 toont de maandelijkse resultaten voor NOD en CBOD van de voorbije 5 jaar. Het is duidelijke dat de zuurstofvraag voor nitrificatie jaarrond nog steeds veel te hoog is (criterium evaluatiemethodiek NOD is 2 mg/l), ook CBOD voldoet vaak niet (criterium CBOD is 6 mg/l). De reden hiervoor is dat meer dan 50% van het debiet van de Zenne afkomstig is van Brusselse RWZIs. Brussel Zuid heeft nog altijd geen tertiaire behandeling waardoor er hoge N vrachten geloosd worden. De zuivering van Brussel Noord heeft wel een efficiënte tertiaire behandeling, maar kwaliteitsnormen voor gezuiverd afvalwater zijn hoger dan voor oppervlaktewater (totaal N= 10 mg/l tegen 2 mg/l in oppervlakte water; BOD = 25 mg/l tegen 6mg/l in oppervlakte water).
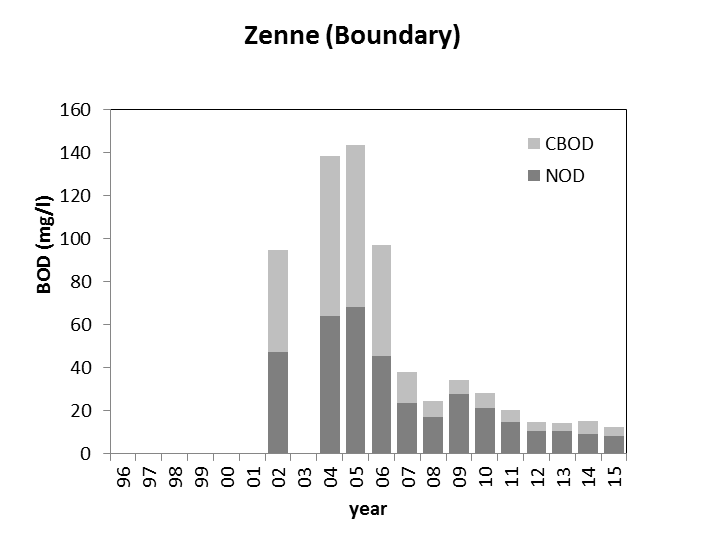
Figuur 4-21 Biochemische zuurstofvraag (BOD) in de Zenne, opgesplitst in zuurstofvraag voor nitrificatie (NOD) en zuurstofvraag voor koolstofmineralisatie (CBOD) (data OMES en VMM).
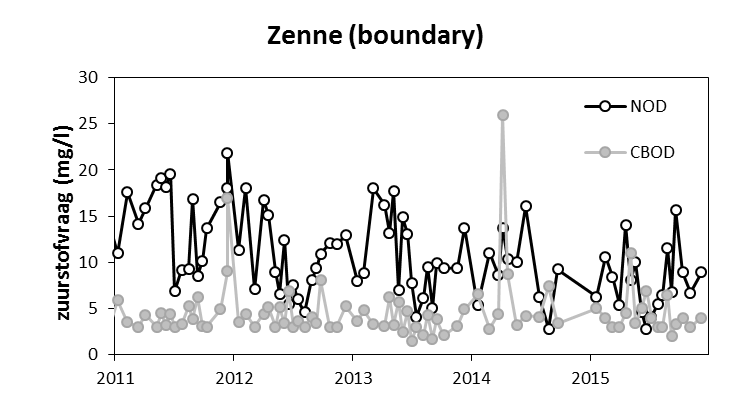
Figuur 4-22 Biochemische zuurstofvraag voor nitrificatie (NOD) en voor koolstofmineralisatie (CBOD) voor de boundary Zenne.
De Rupel kreeg voor de zuivering van Brussel een zeer grote BOD vracht te verwerken, met een groot aandeel NOD. In tegenstelling tot de zoete Zeeschelde, droeg de Rupel zelf waarschijnlijk weinig bij tot de verwerking van die NOD. Ook voor het station Boom op de Rupel werd de theoretisch berekende NODth (op basis van NH4 en NO2) vergeleken met de op metingen van BODwithout en CBOD gebaseerde NODmeas. In de Rupel blijken deze niet gecorreleerd te zijn, ook niet bij temperaturen boven 15°C (Figuur 4-23). Bij temperaturen boven 15 °C, waarbij men zoals in de zoete Zeeschelde een actieve populatie nitrificeerder zou kunnen verwachten, is het verschil tussen BODwithout en CBOD klein. Meestal zijn de metingen met en zonder nitrificatieremmer aan elkaar gewaagd, wat erop wijst dat er geen actieve populatie nitrificeerders aanwezig was in Boom (periode 1998-2003), zelfs niet bij temperaturen boven 15°C, en dit ondanks de aanwezigheid van een hoge ammoniumconcentratie.
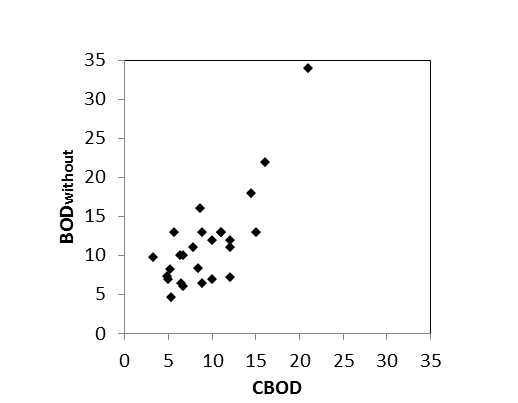
Figuur 4-23 correlatie tussen CBOD en BODwithout bij temperaturen boven 15°C aan station Boom
Voorwaarden voor een actieve populatie nitrificerende bacteriën zijn o.a. een voldoende hoge temperatuur (> 15°C), voldoende zuurstof en ammonium en een voldoende lange verblijftijd. In Dendermonde was deze populatie duidelijk aanwezig, in Boom niet, hoewel de zuurstofwaarden in de periode 1998-2007 van dezelfde grootteorde waren als deze te Dendermonde in de periode 1996-2000. Te Dendermonde zijn ammonium en zuurstof negatief gecorreleerd (Figuur 4-24). In Boom daarentegen tekenen zich twee perioden af: voor de zuivering van Brussel waarbij ammonium hoog was en zuurstof laag, en na de zuivering, met lagere ammonium en een brede range aan hogere zuurstofwaarden. Zuurstof is niet gecorreleerd aan de ammoniumconcentratie en onder goede zuurstofcondities (>5 mg/l) komt nog steeds redelijk veel ammonium voor (>0.5 mg/l), wat er op wijst dat ook na de inwerkingtreding van de zuivering in Brussel, de Rupel geen sterke nitrificatie kent.
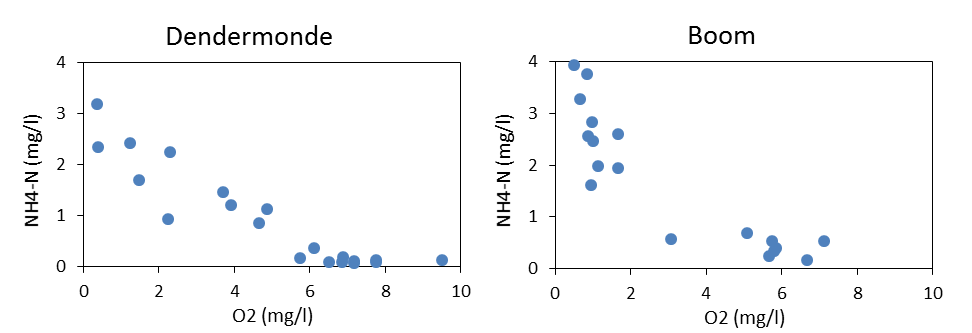
Figuur 4-24 Correlatie tussen zuurstof en ammonium, bij watertemperaturen boven 15°C, voor Dendermonde en Boom.
Van nitrificerende bacteriën is geweten dat ze traag groeien. Voor RWZI’s wordt gesteld dat de verblijftijd van het actief slib (en dus de nitrificerende bacteriën) minstens 4 dagen moet zijn. In de zoete zone is deze hydraulische verblijftijd in de zomer ruim 4 dagen (gemiddeld 9 dagen over traject Melle – Temse). De verblijftijd van het slib ligt, dankzij tidal pumping, wellicht nog hoger. In de Rupel ligt de hydraulische verblijftijd lager: over de ganse Rupel is het zomergemiddelde net geen 4 dagen, zodat halverwege de Rupel, te Boom, de verblijftijd misschien te kort is voor de opbouw van een actieve populatie nitrificeerders.
4.2.Nutriënten
4.2.1.Inleiding
In de Evaluatiemethodiek wordt de toetsparameter nutriënten beoordeeld aan de hand van volgende rekenparameters (Tabel 4-2):
Tabel 4-2 Rekenparameters voor de toetsparameter Nutriënten, volgens de Evaluatiemethodiek Schelde-estuarium
| Reken-parameter | Beoordeling Zeeschelde |
| NOD | maximum 2mg/l, te bepalen in elk Scheldecompartiment |
| CBOD | maximum 6 mg/l niet overschrijden, te bepalen in elk Scheldecompartiment |
| TDIN | mag niet stijgen ten opzichte van de gemiddelde concentratie van de voorbije 6 jaar, te bepalen in elke Scheldezone |
| Fosfaat | mag niet stijgen ten opzichte van de gemiddelde concentratie van de voorbije 6 jaar, te bepalen in elke Scheldezone |
| DSi | Mag niet meer dan 2 opeenvolgende meetcampagnes onder 0,01 mM (= 0,28 mg/l Si) dalen, te bepalen in elk compartiment |
| ICEP-TSi | Deze parameters vereisen fluxberekeningen over het ganse estuarium en kunnen bijgevolg niet gepaald worden in het kader van OMES. |
| ICEP-DSi | |
| TDINflux | |
| Nitriet | maximum 0.1 mg/l NO2--N, te bepalen in elk Scheldecompartiment |
| Ammoniak | maximum 0.025 mg/l NH3-N, te bepalen in elk Scheldecompartiment |
De parameters voor biochemische zuurstofvraag NOD en CBOD werden in vorig hoofdstuk reeds besproken en worden daarom niet herhaald. Wel wordt de evaluatie mee opgenomen in de eindtabel (Tabel 4-3).
4.2.2.Stikstof
Stikstof is, net als fosfor, een essentieel nutriënt voor de groei van algen. N is echter in de Zeeschelde in zo’n grote overmaat aanwezig dat het kan leiden tot buitensporige bloei met potentieel negatieve gevolgen: eutrofiëring. Toch komt het in de Zeeschelde niet tot een ongeremde bloei: de slechte waterkwaliteit in de jaren ’90 en het slechte lichtklimaat houdt algenbloei binnen de perken, al blijft het gevaar wel latent aanwezig. Bovendien stromen nu veel te grote vrachten aan stikstof de Schelde door, die momenteel in het estuarium zelf geen al te grote problemen veroorzaken, maar wel in de kustzone de eutrofiëring voeden. Daarom legt MONEOS een maximum export van totaal anorganische stikstof (TDIN = NH4+ + NO3- + NO2-) vanuit het estuarium naar de kust op, en mag voor een positieve evaluatie de gemiddelde TDIN concentratie niet stijgen ten opzichte van het gemiddelde van de voorbije 6 jaar. Hieraan wordt voldaan (Tabel 4-3): TDIN is nog steeds, zij het beperkt, in dalende lijn (Figuur 4-25).
Figuur 4-25 Gemiddelde TDIN concentratie langsheen de Schelde en Rupel (zwart omlijnde bolletjes). Voor de Rupel (monding op km 92 in de Schelde) is een gemiddelde waarde over de ganse waterloop weergegeven.
De samenstelling van TDIN is de voorbije jaren duidelijk veranderd (Figuur 4-26). In de jaren ’90 vormde ammonium nog een belangrijk aandeel in de totale stikstofconcentratie. Door de sterke toename van het zuurstofgehalte kon dit ammonium efficiënter genitrificeerd worden, waardoor NH4+ daalde, maar NO3- steeg (Figuur 4-28, Figuur 4-29). Door de hogere zuurstofwaarden werd bovendien het pelagiale denitrificatieproces, een anoxisch proces waarbij stikstof als N2-gas verdwijnt uit de waterkolom, sterk geïnhibeerd. De sterke seizoenaliteit waarbij NO3 ‘s zomers sterk daalt door een sterke denitrificatie, nam af. Nitraat is toegenomen in de zomer, afgenomen in de winter.
Stikstof is in dalende lijn in het estuarium, een daling die sneller lijkt te gaan in de Zeeschelde dan in Bovenschelde of Rupel (Figuur 4-26, Figuur 4-27). In Rupel en vooral Zenne is een duidelijke sprong in de N-concentratie te zien in 2007: de waterzuivering speelt daar duidelijk een rol. In de Zeeschelde is de link tussen toenemende capaciteit aan waterzuivering en de daling in TDIN veel minder duidelijk (Figuur 4-34): TDIN daalt quasi lineair in de tijd, wat erop kan wijzen dat de bijdrage aan N-verwijdering door RWZI’s beperkt is. De daling wordt vermoedelijk meer veroorzaakt door wijzigingen in het landbouwbeleid. Struyf et al (2004) hadden eerder al aangetoond voor de Zeeschelde dat de TDIN vracht niet zozeer van puntbronnen (zoals afvalwater lozingen) komt, maar eerder diffuus is.
Voor ammonium en nitraat legt de Evaluatiemethodiek geen maxima op, maar een maximum voor ammonium zit impliciet vervat in de NOD, aangezien het in hoofdzaak NH4+ is die verantwoordelijk is voor de zuurstofvraag voor nitrificatie.
Omwille van de toxiciteit van ammoniak en nitriet, zijn voor deze stikstofvormen wel criteria opgenomen: maximaal 0,025 mg/l voor NH3 en 0,1 voor NO2-. Op Figuur 4-33 is duidelijk te zien dat nitriet in dalende lijn is, maar dat anno 2015 in de zoete zone nog niet wordt voldaan aan de criteria uit de Evaluatiemethodiek. Na de sterke daling in de periode 2000 – 2010 wordt recent ook geen significante afname meer geconstateerd en blijft vooral de zoete Zeeschelde nog te veel nitriet (meer dan 0,1 mg/l) bevatten. Enkel de zone met sterke saliniteitsgradiënt voldoet (Tabel 4-3).
In de jaren ’90 kwam ook het toxische ammoniak in te hoge concentraties voor in de Zeeschelde (Figuur 4-32). Na de shift in 2003 nam ammonium zeer sterk af, zodat ook het voorkomen van ammoniak sterk daalde. NH3 bleef dan beperkt tot de zoete zone met korte verblijftijd. Na 2008, wanneer de zoete zone steevast hogere zuurstofwaarden kent en NH4 enkel in lage concentraties voorkomt, verdwijnt NH3 uit de Zeeschelde, ondanks een sterke stijging van de pH (Figuur 4-43). Een hoge pH duwt immers het chemische evenwicht van NH4+ richting NH3, maar de NH4+-concentraties zijn laag genoeg om ammoniak toxiciteit te vermijden. In 2015 is de evaluatie voor deze parameter daarom positief (Tabel 4-3).
De organische fractie schommelt sterk met de seizoenen, maar kent jaargemiddeld geen duidelijke af- of toename (Figuur 4-27, Figuur 4-31). Vermoedelijk is er wel, net als bij koolstof, een verschuiving van allochtoon naar meer autochtoon materiaal.
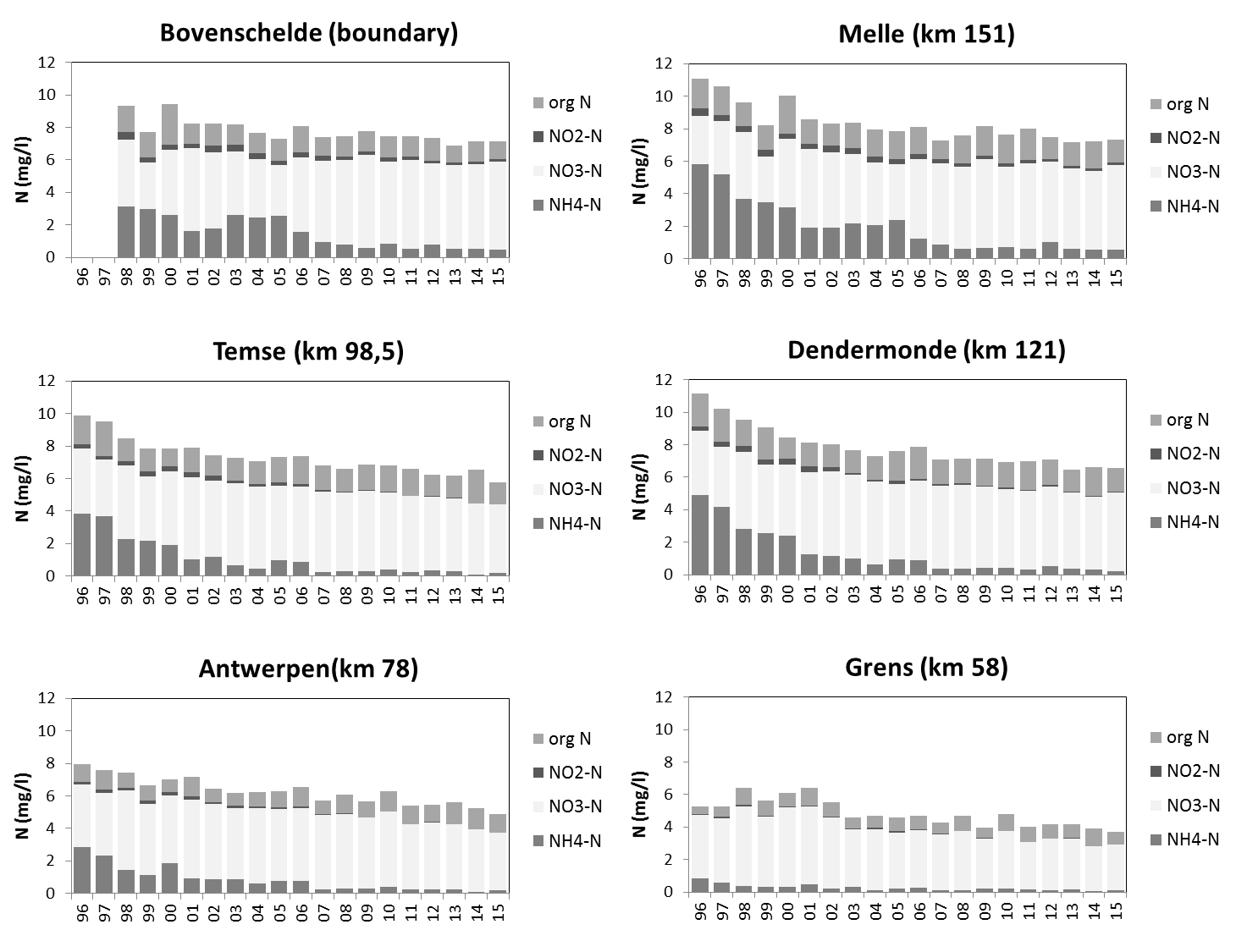
Figuur 4-26 Stikstofconcentratie, opgedeeld in de fracties organische stikstof (OrgN), nitriet (NO2-N), nitraat (NO3-N) en ammonium (NH4-N), voor diverse stations.
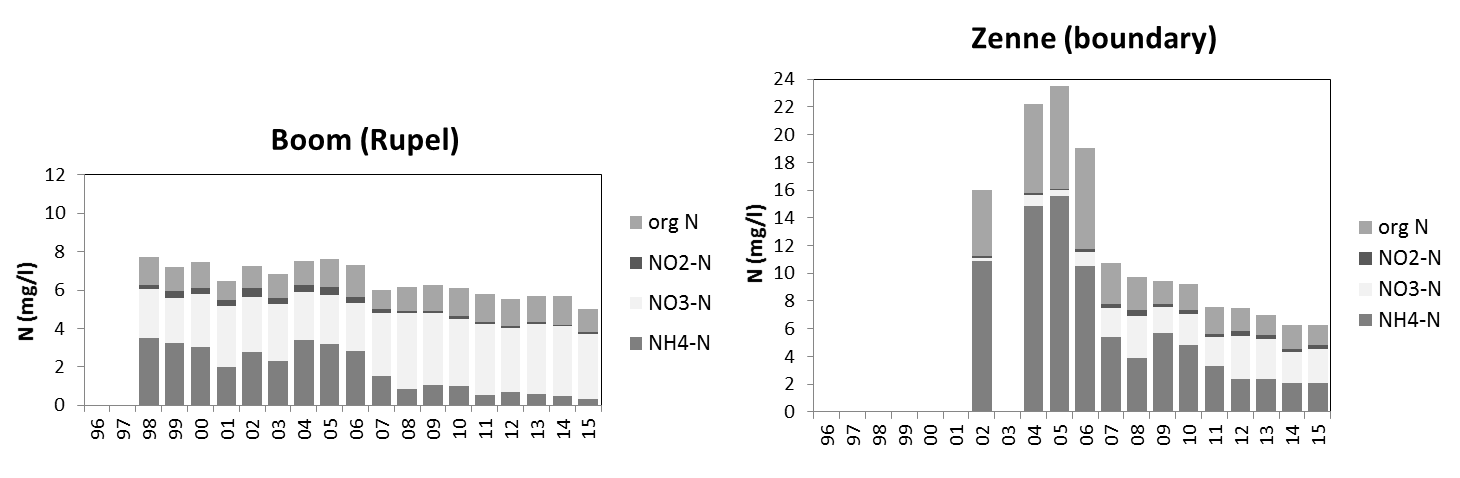
Figuur 4-27 Stikstofconcentratie, opgedeeld in de fracties organische stikstof (OrgN), nitrite (NO2-N), nitraat (NO3-N) en ammonium (NH4-N), voor het station Boom en de boundary Zenne.
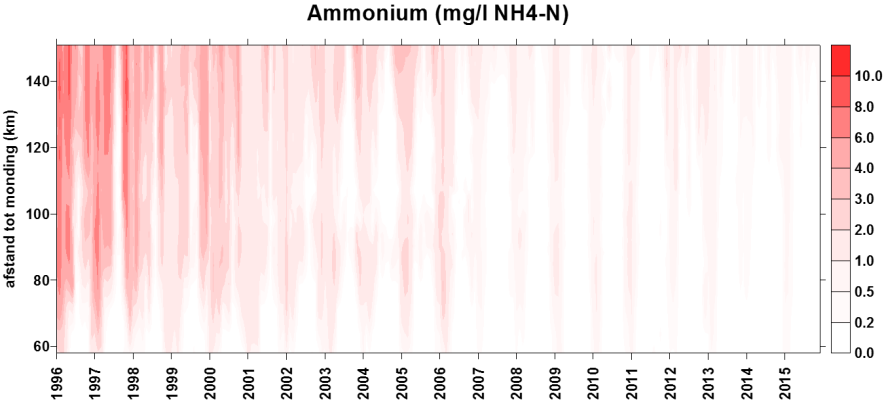
Figuur 4-28 Surface plot voor ammonium.
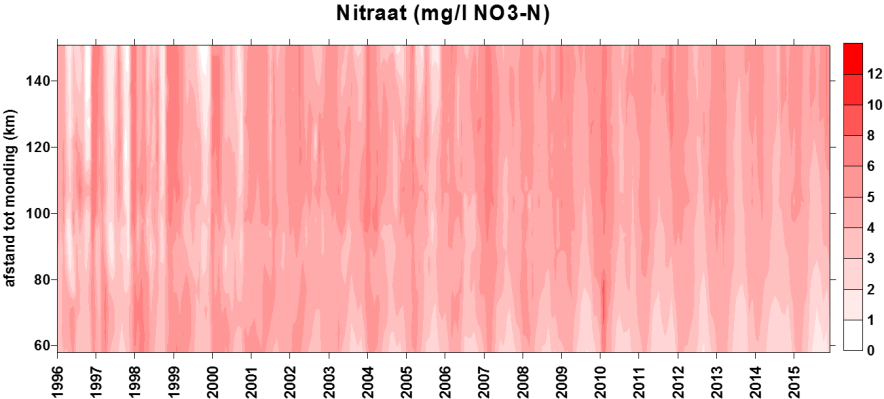
Figuur 4-29 Surface plot voor nitraat.
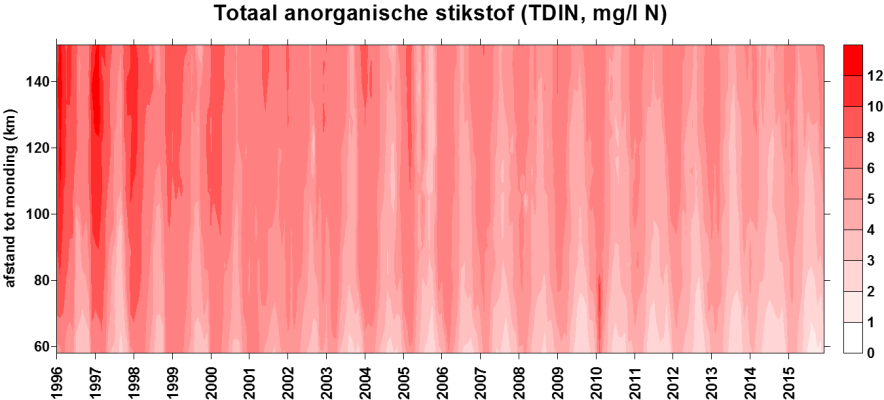
Figuur 4-30 Surface plot voor totaal anorganische stikstof (TDIN = NH4-N + NO3-N + NO2-N).
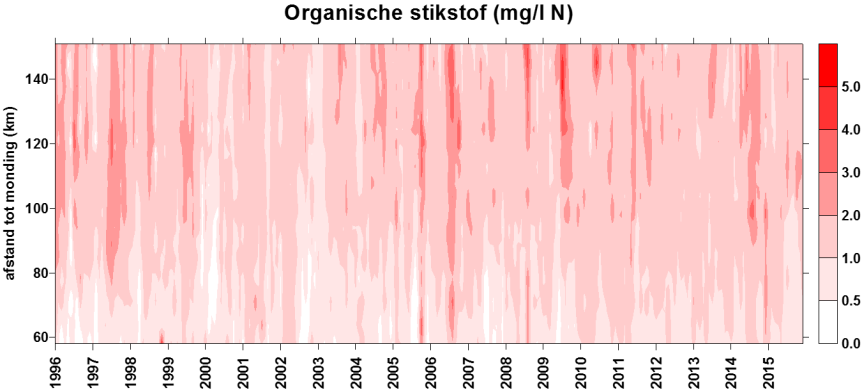
Figuur 4-31 Surface plot voor organische stikstof.

Figuur 4-32 Surface plot voor ammoniak (berekend). De zwarte lijn markeert de grens van 0.025 mg/l (Evaluatiemethodiek)
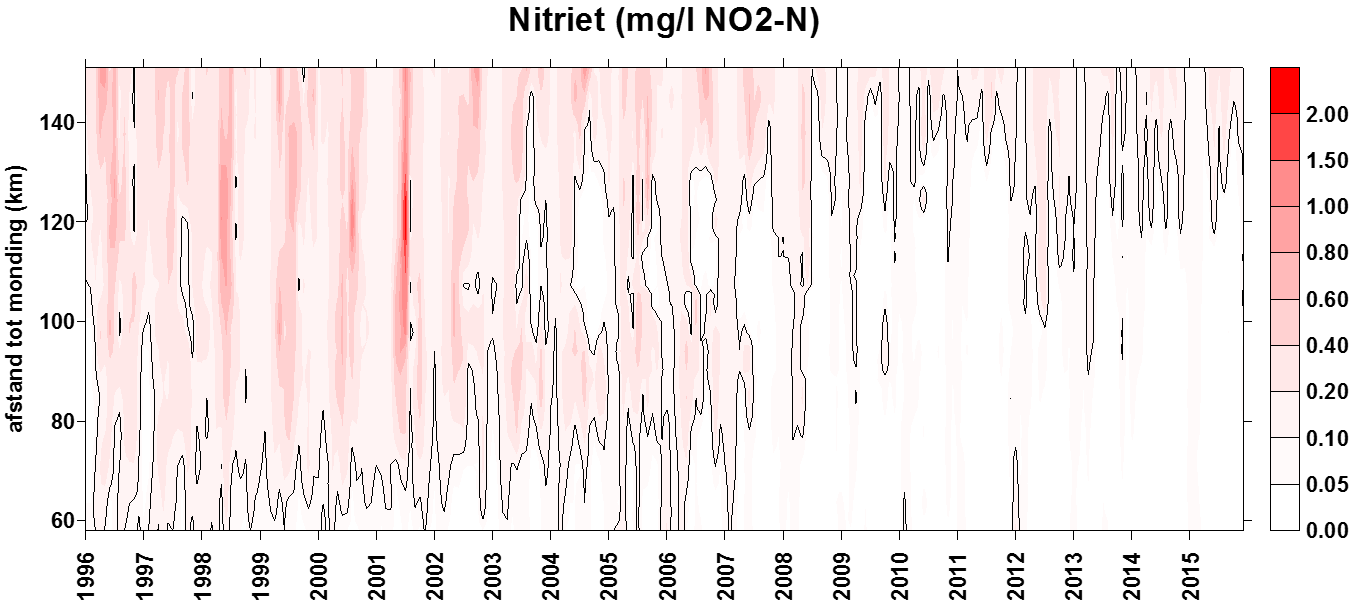
Figuur 4-33 Surface plot voor nitriet, met in zwart de 0.1 mg/l grens (Evaluatiemethodiek)
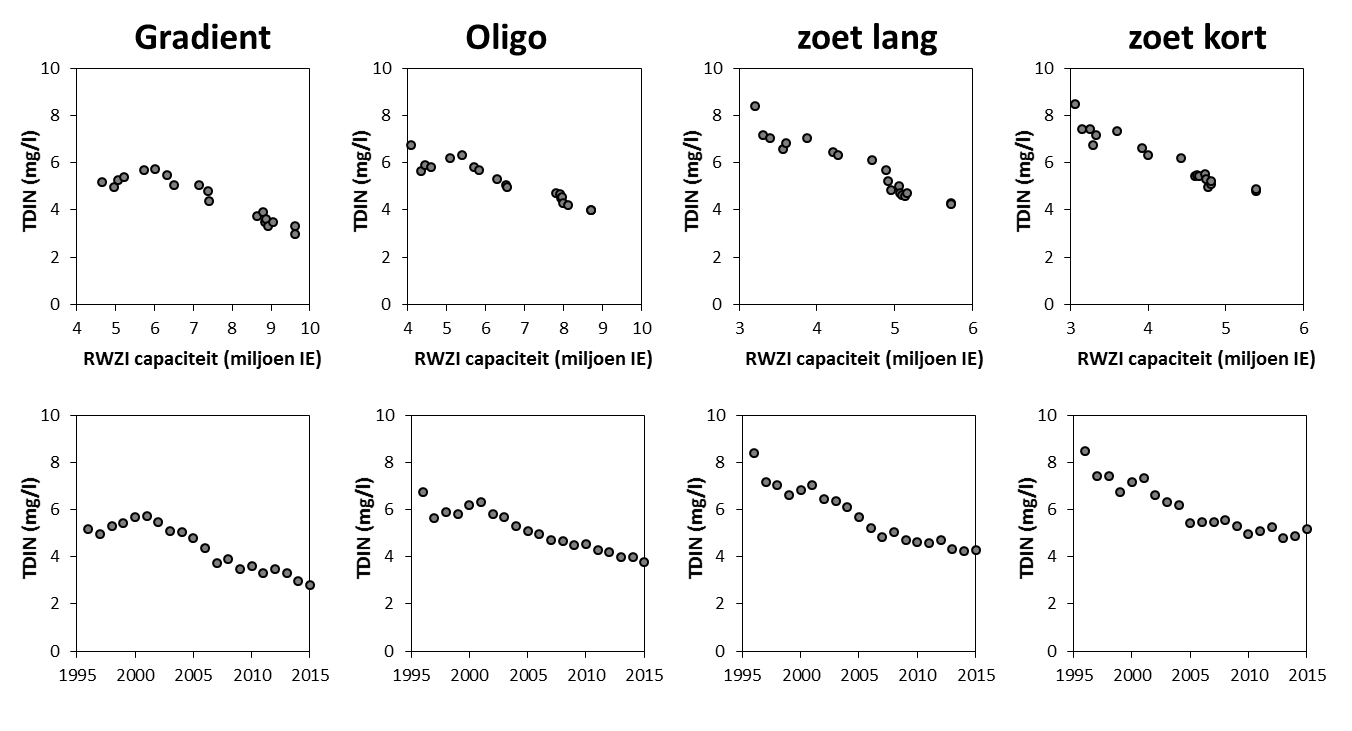
Figuur 4-34 TDIN (mg/l) per zone in het zomerhalfjaar in functie van de rioolwaterzuiveringscapaciteit (RWZI capaciteit, in miljoen inwoner equivalenten (IE)), en in functie van de tijd.
4.2.3.Fosfor
Hoewel minder abundant aanwezig dan stikstof, is ook fosfor doorgaans in veel te hoge concentratie aanwezig in het Scheldewater. De Evaluatiemethodiek stelde daarom dat de gemiddelde fosfaatconcentratie niet mag toenemen ten opzichte van het gemiddelde van de voorbije zes jaar. Uit Figuur 4-35 blijkt dat fosfaat een sterke daling kende ten opzichte van 20 jaar geleden, maar dat fosfaat de laatste jaren nog amper daalt. In 2015 was er in de zoete zone met kort verblijftijd zelf een beperkte toename van fosfaat ten opzichte van de voorbije zes jaar, wat een negatieve evaluatie oplevert voor deze zone (Tabel 4-3). Let wel, de Evaluatiemethodiek beschouwt gemiddelden over zes jaar, in deze jaarlijkse OMES rapportage wordt voor 2015 een jaargemiddelde vergeleken met het gemiddelde van de zes voorgaande jaren).
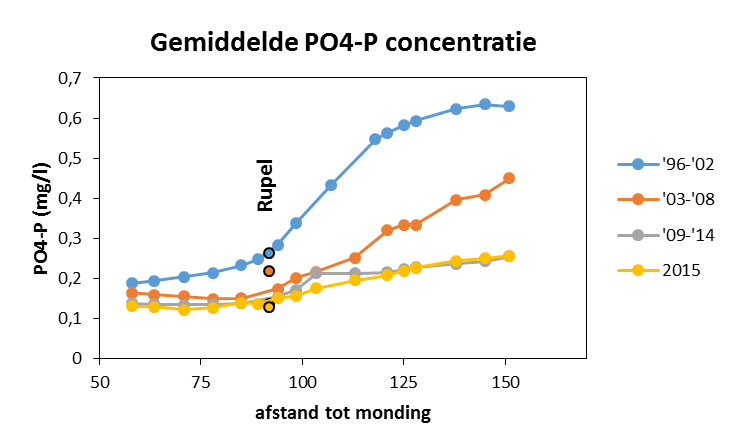
Figuur 4-35 Gemiddelde fosfaat concentratie langsheen de Schelde en Rupel (zwart omlijnde bolletjes). Voor de Rupel (monding op km 92 in de Schelde) is een gemiddelde waarde over de ganse waterloop weergegeven.
De surface plot voor fosfaat (Figuur 4-38) illustreert mooi de sterke afname van fosfaat. De sterkste afname situeert zich in de zoete zones eind jaren ’90, en wordt mee verklaard door de sterke afname in de Bovenschelde. Nadien neemt de daling af en vanaf 2010 blijft fosfaat ongeveer constant.
Een lange termijn plot aan station Grens (Figuur 4-36) toont hoe fosfaat rond 1980 een maximum bereikte, om vanaf dan zeer sterk af te nemen. Aan de grens vertraagt die daling vanaf 1995, maar zet zich wel continu voort tot in 2015. De sterke en abrupte daling van de jaren ’80 kan niet verklaard worden door toenemende RWZI capaciteit; deze nam amper toe in die periode (Figuur 4-36). De omschakeling naar meer fosfaatvrije waspoeders vanaf eind jaren ’80 ligt hier wellicht aan de basis.
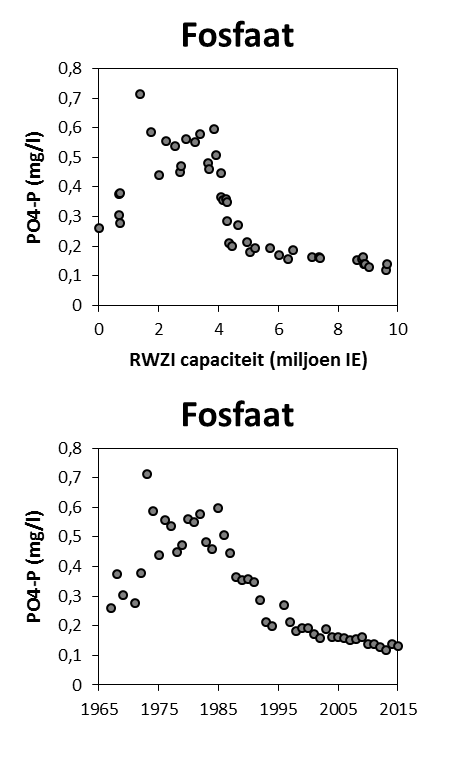
Figuur 4-36 Jaargemiddelde fosfaatconcentratie (mg/l) aan het station Grens, in functie van de rioolwaterzuiveringscapaciteit (RWZI capaciteit, in miljoen inwoner equivalenten (IE)), en in functie van de tijd.
Ook totaal fosfor (Ptot, Figuur 4-39) kent een afname, maar die beperkt zich vooral tot de jaren ’90. Nadien is er weinig tot geen afname meer, hoewel fosfor nog wel verder daalde. Een mogelijke verklaring, is een toegenomen binding van fosfor, hetzij in biota, hetzij aan partikels. Fosfaten kunnen immers gebonden worden aan sediment en zwevende stof via metaal oxyhydroxides onder gunstige zuurstofomstandigheden. Sinds 2003 zijn die gunstige zuurstofomstandigheden aanwezig in het zoete. Fluctuaties in zwevende stof kunnen mee aan de basis liggen van het sterk schommelende Ptot patroon. De hoogste Ptot waarden zijn doorgaans te vinden rond km 100 nabij het turbiditeitsmaximum.
Niet enkel de absolute concentraties, maar ook de onderlinge verhouding tussen nutriënten is van belang. Dit zal bepalen welk element limiterend kan worden. Algemeen worden N en P in de verhouding 16/1 opgenomen door de algenpopulatie. De N/P ratio vertoont in het ganse estuarium een duidelijk seizoenaal patroon, met maxima in de winter, minima in de zomer. Met andere woorden in de zomer is, relatief ten opzichte van P, N minder abundant aanwezig (Figuur 4-37). In de zomer ligt de N concentratie een stuk lager door bacteriële verwijdering en N opname door algen. P kent een veel kleinere seizoenale schommeling. In de oligohaliene (Temse) en gradiëntzone (Grens en Antwerpen) is N steeds het meest abundante element (N/P ratio > 16). Tot 2006 werd in de zomer in de zoete zone (Dendermonde) P occasioneel het meest abundant. De N/P ratio is echter in stijgende lijn in het zoete. Vanaf 2006 is N overal in het estuarium het meest abundant. Relatief gezien is de P concentratie sterker afgenomen dan de N concentratie.
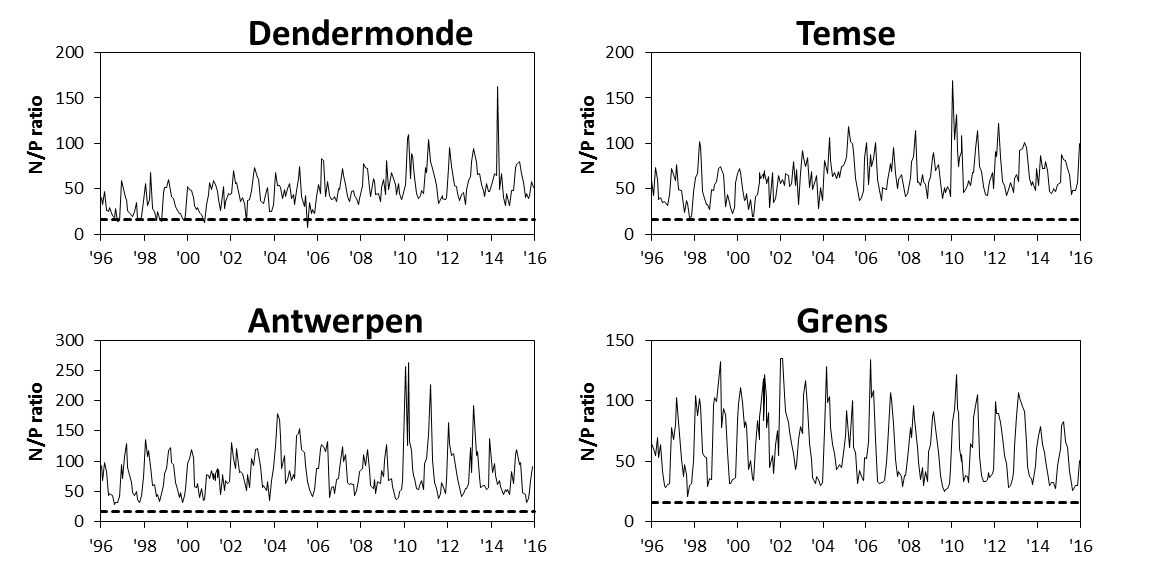
Figuur 4-37 Molaire N/P ratio voor Antwerpen, Temse, Dendermonde en Melle. De streeplijn duidt de ideale N/P ratio van 16/1 aan.
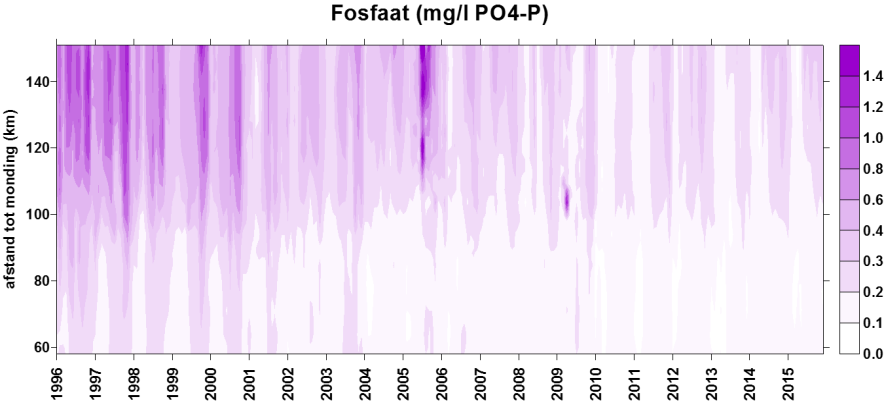
Figuur 4-38 Surface plot voor fosfaat
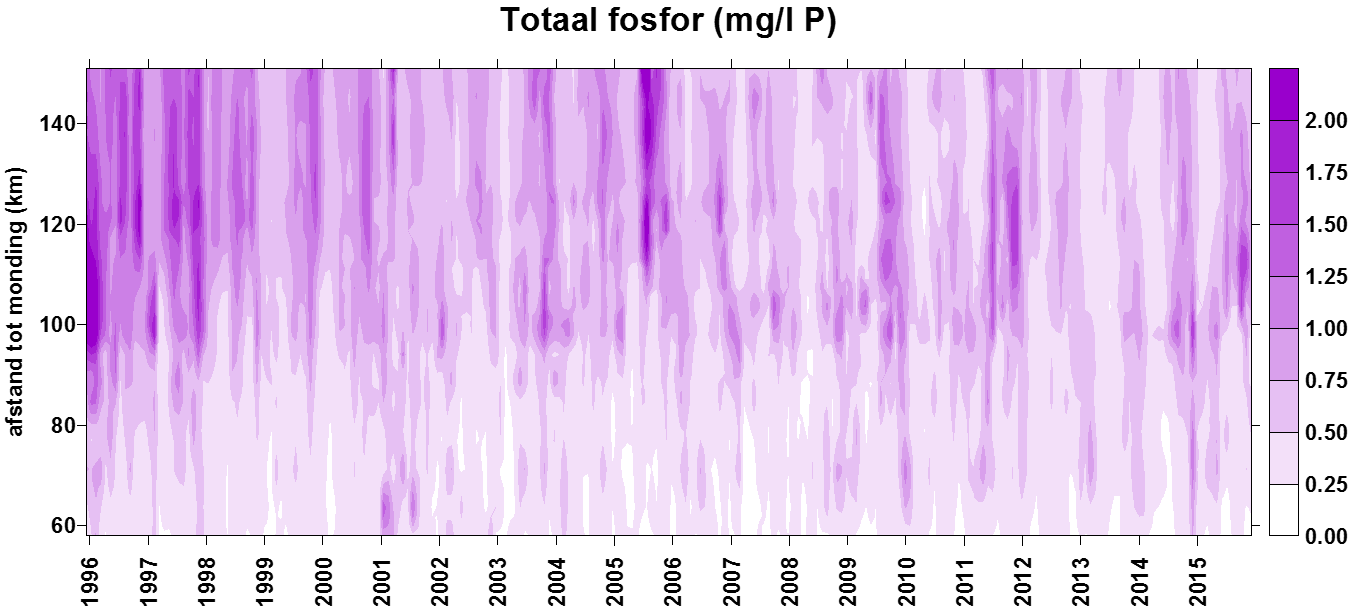
Figuur 4-39 Surface plot voor totaal fosfor
4.2.4.Silicium
Naast stikstof en fosfor krijgt het element silicium bijzondere aandacht als essentieel voedingselement voor diatomeeën. Diatomeeën nemen opgelost silicium (DSi) op voor de bouw van hun skelet. De aanwezigheid van DSi is dus essentieel voor de groei van diatomeeën. Ook N en P zijn essentieel, maar doorgaans zijn deze nutriënten, in tegenstelling tot DSi, in overvloed aanwezig in het Scheldewater. Bij gebrek aan DSi, zullen gewone groenalgen die geen skelet en dus geen silicium nodig hebben, het overnemen van de diatomeeën. Echter, deze groenalgen zijn minder geschikt als voedsel, waardoor de voedselketen wordt verstoord. Daarom stelt de Evaluatiemethodiek dat perioden van siliciumdepletie (minder dan 0.01 mM of 0.28 mg/l) niet te lang mogen duren. Meer dan twee opeenvolgende campagnes met siliciumtekort leiden tot een negatieve evaluatie.
Perioden van sterke diatomeebloei tekenen zich duidelijk af in het DSi verloop: tijdens deze algenbloei dalen de DSi concentraties sterk (Figuur 4-41, Figuur 4-40). Tijdens de zeer sterke algenbloei in de zomer van 2004 bijvoorbeeld, was er een lange periode van zeer lage DSi waarden, met een groot risico op schadelijke groenalgenbloei.
De laatste jaren is de zomerse DSi daling iets minder uitgesproken dan een tiental jaar geleden. Jaargemiddeld lag de chl a concentratie soms iets lager, maar toch komen hoge pieken in algenbloei en dus sterke DSi consumptie nog veelvuldig voor. Maar de duur van de bloei is nu eerder beperkt in de tijd, wellicht ten gevolge van begrazing door de sterk toegenomen zoöplanktongemeenschap. Mogelijks zorgt een efficiënte begrazing ook voor het snel terug vrijstellen van DSi, zodat DSi tekorten worden vermeden. In 2015 kwamen geregeld siliciumtekorten (Figuur 4-40) voor, doch echter steeds van korte duur zodat ze geen problemen veroorzaakten. De rekenparameter DSi krijgt daarom een positieve evaluatie voor de gehele Zeeschelde (Tabel 4-3).
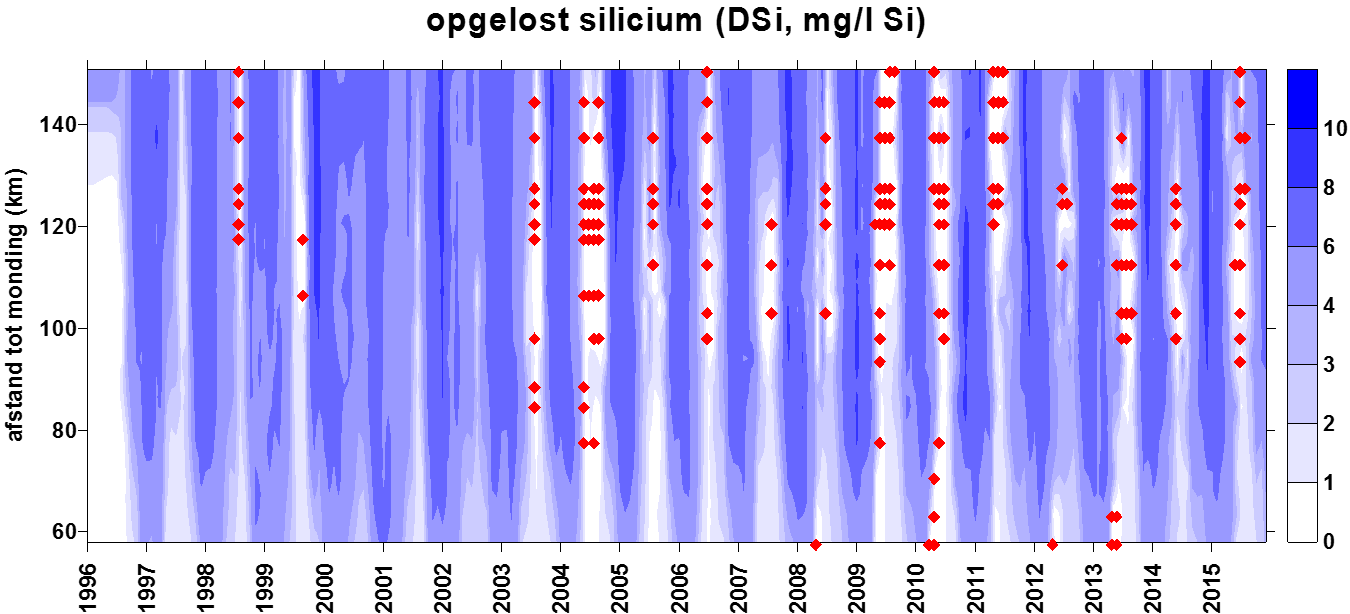
Figuur 4-40 surface plot voor opgelost silicium (DSi). De rode stippen geven concentraties weer onder 0.01 mM, de grens voor siliciumdepletie.
Als silicium wordt opgenomen door organismen en ingebouwd in biologische structuren, spreken we van biogeen silicium (BSi). Dit BSi is niet meer rechtstreeks opneembaar door algen, maar moet eerst terug afgebroken worden tot DSi. BSi wordt in de Schelde sinds 2008 systematisch mee gemeten (Figuur 4-41). Op deze figuren is duidelijk de interactie tussen algenbloei (chl a) en DSi en BSi te zien. Bij algenbloei wordt DSi door diatomeeën geconsumeerd en omgezet in BSi. De pieken in chl a vallen mooi samen met pieken in BSi. Figuur 4-42 toont ter illustratie de correlatie tussen Chl a en BSi voor verschillende perioden. Voor bijvoorbeeld augustus 2013 is een duidelijk lineair verband zichtbaar, dat illustreert dat het aanwezige BSi in de Schelde afkomstig is van de aanwezige algenpopulatie. Over een ruimere periode beschouwd, is dat verband minder duidelijk: het BSi gehalte verschilt wellicht sterk van algensoort tot algensoort, en de verhouding aan soorten is geen constante in de Schelde. Ook het aandeel afgestorven fytoplankton, wat BSi bevat maar geen chl a meer, kan sterk variëren.
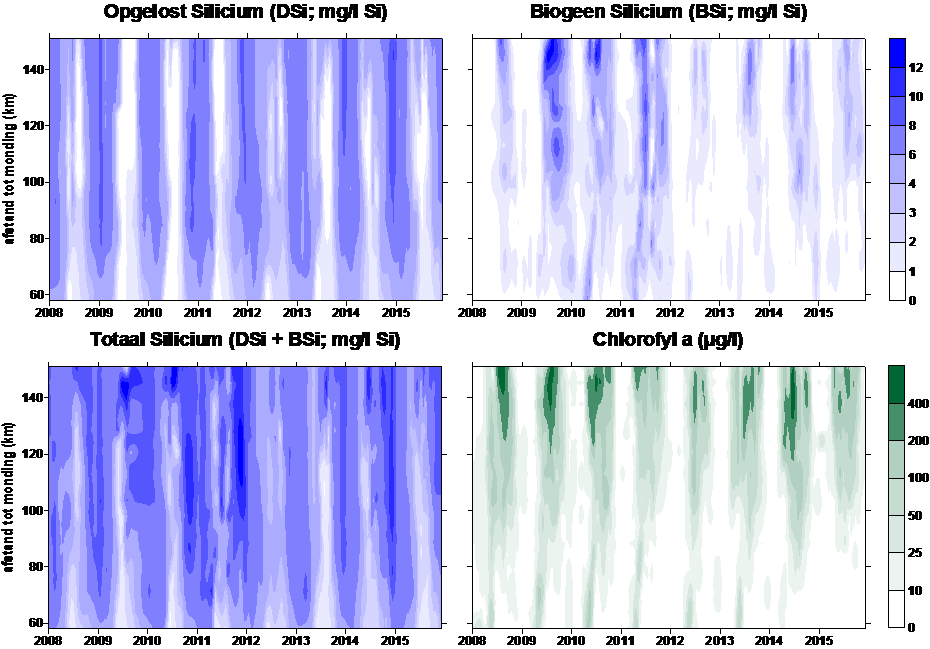
Figuur 4-41 surface plots voor opgelost silicium (DSi), biogeen silicium (BSi), totaal silicium (DSi + BSi) en chl a.
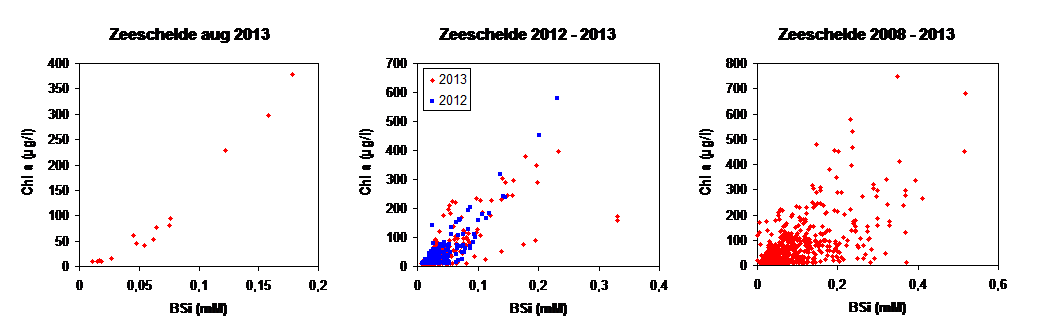
Figuur 4-42 correlatie tussen BSi en chl a in de Zeeschelde
Tabel 4-3 Samenvattende tabel voor de evaluatie van de toetsparameter Nutriënten voor 2015. Voor NOD, CBOD, NO2-N en NH3-N bevat de tabel de maximale waarden (mg/l). De rood gemarkeerde data overschrijden het criterium dat boven elke kolom staat vermeld. Voor TDIN en PO4-P is telkens een jaargemiddelde concentratie weergegeven. Deze mag niet stijgen ten opzichte van het gemiddelde van de voorbije 6 jaar. Voor Si zijn de minima weergegeven. Er volgt pas een negatieve evaluatie als meer dan twee opeenvolgende campagnes Si onder het minimum (0,28 mg/l) duikt.
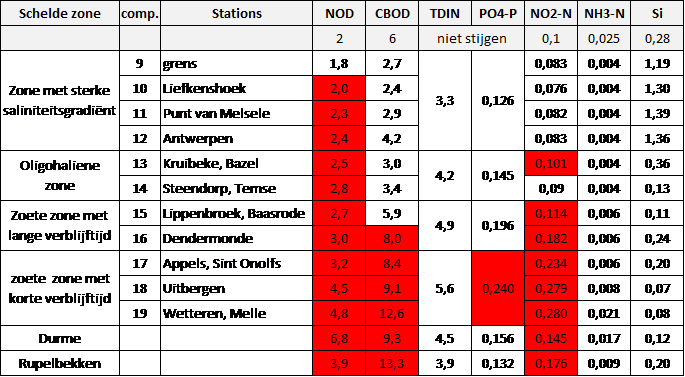
De evaluatie voor verschillende rekenparameters uit de toetsparameter Nutriënten, volgens de Evaluatiemethodiek Schelde-estuarium, wordt samengevat in Tabel 4-3. De beoordelingscriteria staan opgelijst in Tabel 4-2, en hier bondig samengevat bovenaan elke kolom. Aangezien de rekenparameters ICEP-DSi, ICEP-TSi en TDINflux een fluxberekening vereisen, valt de bepaling van deze parameters buiten het bestek van OMES en zijn ze hier niet opgenomen.
De samenvattende tabel bevat nog veel rode cijfers: de waterkwaliteit in de Zeeschelde voldoet duidelijk nog niet aan de vereisten voor de goede basiskwaliteit die de Evaluatiemethodiek nastreeft. Nabij de grens voldoet het Scheldewater echter wel aan alle beschouwde criteria voor de toetsparameter Nutriënten. Dankzij het herstellende ecosysteem wordt de vuilvracht blijkbaar voldoende verwerkt voordat ze de Zeeschelde verlaat richting Westerschelde.
4.3.Zuurtegraad (pH)
De omschakeling van een respiratie gedomineerd systeem naar een primaire productie gedomineerd systeem uit zich ook duidelijk in de sterk gestegen pH (Figuur 4-43). Sterke respiratie zorgde immers voor verzuring, primaire productie doet pH daarentegen stijgen. Een gedaalde respiratie en een sterke primaire productie hebben de pH in de Zeeschelde met ongeveer 1 eenheid verhoogd de voorbije 20 jaar! De stijgende trend lijkt zich nog steeds door te zetten.
Figuur 4-43 Surface plot voor de zuurtegraad (pH) in de Zeeschelde.
Hoofdstuk 5.Lichtklimaat en zwevende stof
5.1.Inleiding
Vermits fotosynthese, primaire productie, afhankelijk is van licht, is de hoeveelheid licht die kan doordringen in de waterkolom van elementair belang voor het ecosysteem. Door het troebele water kan slechts een beperkte fractie van het invallende zonlicht doordringen in de waterkolom in de Zeeschelde, en vormt lichtbeschikbaarheid de limiterende factor voor algenbloei. Algenbloei, en bijgevolg ook waterkwaliteit en het functioneren van het ganse ecosysteem zijn dus in belangrijke mate afhankelijk van dit lichtklimaat. Daarom worden binnen OMES het lichtklimaat en de factoren die het lichtklimaat beïnvloeden nauwkeurig opgevolgd.
Figuur 5-1 toont schematisch de eufotische diepte. Dit is de diepte tot waar het invallende licht kan doordringen, gedefinieerd als de diepte waarop nog slechts 1% van het invallende licht aanwezig is. Dit wordt beschouwd als de diepte tot waar algen aan fotosynthese kunnen doen.
In een estuarium zoals de Schelde wordt de waterkolom goed gemengd. De algen zijn hierdoor steeds in beweging en maar een korte periode in de eufotische zone, en veel langer in de diepere, donkere waterlagen (Figuur 5-1). Daarom wordt het lichtklimaat niet enkel bepaald door de eufotische diepte (Zeu) , maar evenzeer door de mengdiepte (Zm). De ratio Zeu/Zm bepaalt of het lichtklimaat voldoende is voor primaire productie. Bij Zeu/Zm > 1 kan het licht doordringen over de ganse waterkolom en ontvangt het fytoplankton zich gedurende de dag altijd licht. Bij Zeu/Zm < 1 wordt enkel het bovenste deel van de waterkolom belicht. Als Zeu/Zm = 0,2 bijvoorbeeld, betekent dit dat de algen 20% van de dag in de bovenste, belichte zone zijn en 80% in het donker.
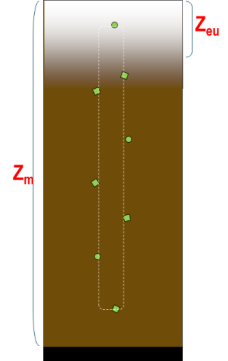
Figuur 5-1 Schematische voorstelling van een alg in de waterkolom met aanduiding van de eufotische diepte (Zeu) en mengdiepte (Zm).
Binnen OMES wordt de eufotische diepte bepaald aan de hand van de lichtextinctiecoëfficiënt kD. De lichtextinctie is afhankelijk van de hoeveelheid zwevende stof in het water (Figuur 5-2). Daarom verdient zwevende stof ook in een rapport over de ecologische kwaliteit van het estuarium alle aandacht. De omes dataset voor zwevende stof start bovendien in december 1995, en vormt daarmee een van de meest coherente datasets om de evolutie van zwevende stof in de Zeeschelde te beschrijven.
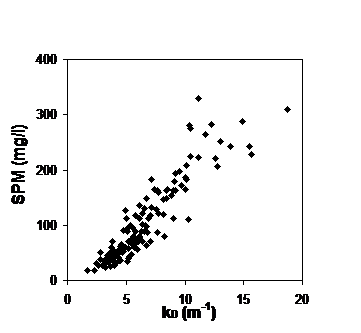
Figuur 5-2 Correlatie tussen de lichtextinctiecoëfficiënt (m-1) en de concentratie zwevende stof (mg/l) nabij het oppervlak
Zwevende stof (SPM) fluctueert echter sterk binnen 1 tijcyclus. Binnen OMES is gekozen om tijonafhankelijk te bemonsteren. Dit impliceert dat de staalname op verschillende momenten in de tijcyclus wordt uitgevoerd, met onvermijdelijk een zekere variatie in zwevende stof tot gevolg. Om deze SPM variatie te kaderen in de tijcyclus, wordt daarom stroomsnelheid en –richting mee geregistreerd. Zwevende stof zal daarom geanalyseerd worden in functie van de stromingskarakteristieken. Door uit te middelen over seizoenen of jaren, kan ook de tijvariatie grotendeels uitgevlakt worden.
In IMDC (2013) werd geverifieerd dat de getij-onafhankelijke meetstrategie geen invloed heeft op het jaar- of seizoensgemiddelde, op basis van een vergelijking tussen de OMES-staalnames op de meetlocatie ‘Melsele’ en metingen van het nabijgelegen permanente meetstation ‘Oosterweel Boven’ die worden uitgevoerd in het kader van MONEOS. In het permanente meetstation wordt om de 10 minuten zwevende stof bepaald via het opmeten van troebelheid (FTU). Op basis van deze 10-minutelijkse tijdreeks werd de jaargemiddelde troebelheden berekend. Daarnaast werd in IMDC (2013) de jaargemiddelden bepaald voor de laagfrequentere OMES tijdreeks nabij Oosterweel (meetlocatie ‘Melsele’). Figuur 6-3 toont aan dat beide tijdreeksen een gelijkaardig patroon vertonen en elkaar benaderen. Hieruit kan besloten worden dat de jaargemiddelden van de periodieke tijdreeksen op zijn minst een goed beeld geven van de jaarlijkse trend over de verschillende meetjaren heen. Deze gemiddeldes zullen gebruikt worden voor het uitvoeren van een analyse van zwevende stof over de jaren heen en de ruimtelijke variatie in het meetgebied.
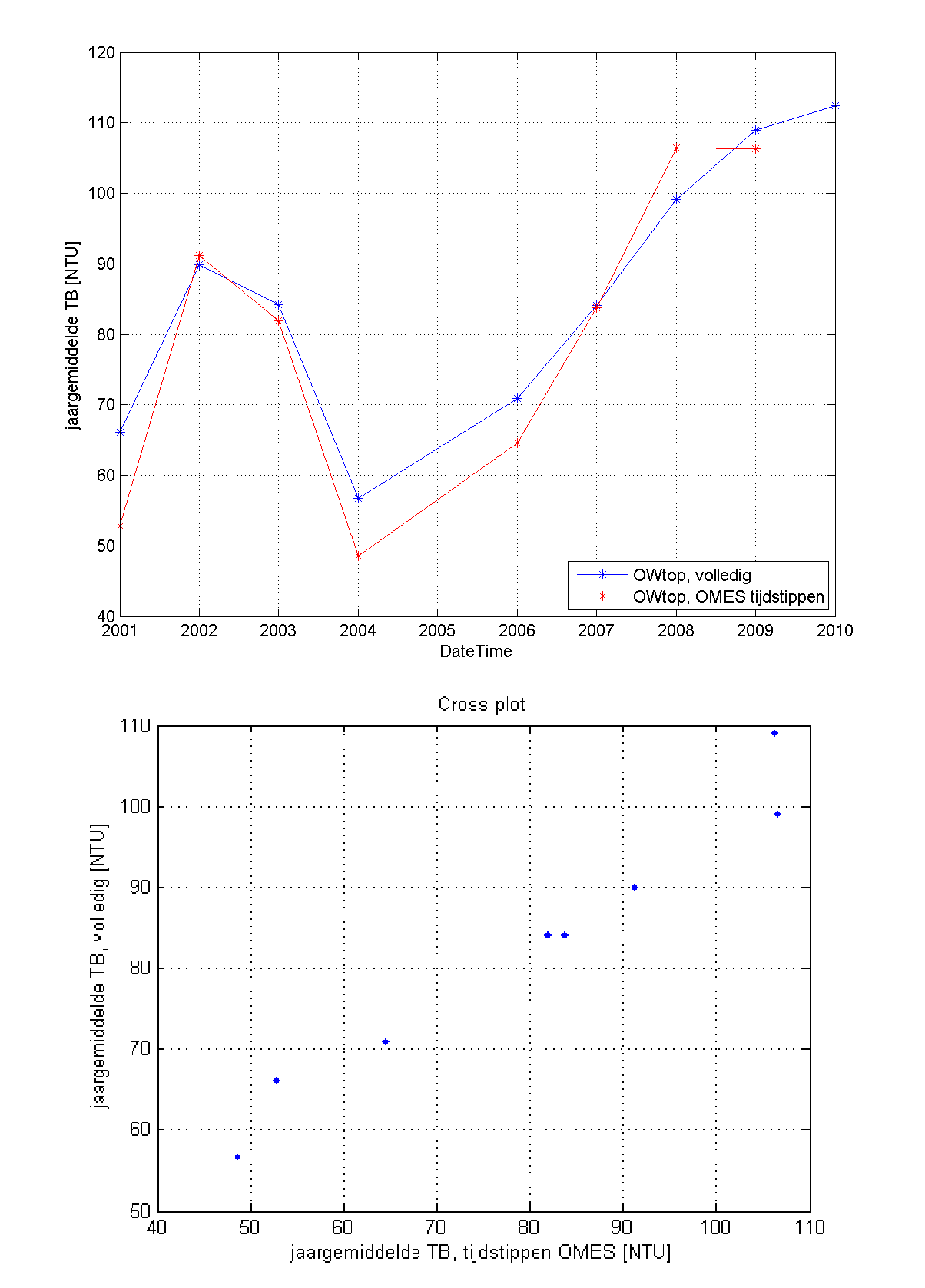
Figuur 5-3: Boven: Jaargemiddelde troebelheid berekend op basis van de hoog- (blauw) en laagfrequente (rood) tijdreeksen te Oosterweel. Onder: Cross-correlatie tussen de jaargemiddelden van de laag- en hoogfrequente tijdreeksen(r = 0.98). (Bron: IMDC, 2013).
In tegenstelling tot de opgeloste stoffen, vertoont zwevende stof een duidelijke dieptegradiënt. Daarom zijn voor zwevende stof verticale profielen opgenomen.
5.2.Getijvariatie
5.2.1.Kruibeke, Terhagen en Schoonaarde
Zwevende stof en de stroomkarakteristieken variëren over een getij; dit wordt geïllustreerd in de 13-uursmetingen te Kruibeke, Terhagen en Schoonaarde. De resultaten van meetjaar 2015 worden gepresenteerd in bijlage en worden verder in deze sectie besproken.
De 13-uursmetingen aan Kruibeke en Schoonaarde werden onder gemiddelde tijcondities uitgevoerd en de meting aan Terhagen onder springtijcondities. De 3 meetlocaties vertonen ongeveer een gelijkaardig snelheidsverloop. De hoogste stroomsnelheden komen voor tijdens de vloedfase ongeveer 1 uur voor HW. Deze stroomsnelheden bereiken te Schoonaarde een dieptegemiddelde waarde van 1.17 m/s en voor Terhagen een waarde van 1.27 m/s. Voor Kruibeke werd een maximum van 0.99 m/s gemeten. Tijdens kentering worden de laagste stroomsnelheden waargenomen; deze vinden plaats ongeveer 30 minuten na HW of LW.
De gemiddelde stroomsnelheden zijn tijdens de ebfase groter dan tijdens de vloedfase aan Kruibeke en Terhagen. Bij Schoonaarde wordt het omgekeerde patroon waargenomen. Hetzelfde patroon werd ook waargenomen in de voorgaande meetjaren vanaf 2012. Uitgezonderd van meetjaar 2011, waar telkens de stroomsnelheid tijdens de vloedfase hoger was dan tijdens de ebfase. In dit meetjaar werden er raaien gevaren in plaats van het nemen van op een vast meetpunt. De hogere gemiddelde snelheid tijdens eb dan tijdens vloed is mogelijk een gevolg van de positie van het vast meetpunt.
Ter hoogte van Kruibeke variëren de waargenomen dieptegemiddelde zwevende stofconcentraties tussen 53 en 220 mg/l. De hoogste dieptegemiddelde concentratie is waargenomen in de vloedfase 1 uur voor HW en de laagste tijdens de LW-kentering. Op deze momenten zijn ook de dieptegemiddelde snelheden het hoogst en het laagst. Tijdens de HW-kentering zijn de concentraties ook laag. De gemiddelde concentraties over de vloed- en ebfase zijn gelijkaardig (ca. 125 mg/l). Deze grootorde van concentraties sluit aan de waargenomen concentraties in 2014 die lager waren dan de voorgaande meetjaren.
Gelijkaardig aan Kruibeke, worden de laagste zwevende stofconcentraties waargenomen bij de kenteringen aan Schoonaarde en Terhagen. Bij beide locaties worden de hoogste concentraties waargenomen rondom 1 uur voor en na LW-kentering. De dieptegemiddelde zwevende stofwaarnemingen aan Schoonaarde en Terhagen variëren tussen dezelfde orde van ca. 50 tot 300 mg/l. Aan Schoonaarde zijn de gemiddelde zwevende stofconcentraties gelijkaardig tijdens eb- en vloedfase (ca. 150 mg/l), maar aan Terhagen is de gemiddelde concentratie tijdens vloed ca. 40 mg/l groter dan tijdens eb (ca. 180 mg/l).
Algemeen kunnen we stellen dat hogere zwevende stofconcentraties voorkomen bij hogere stroomsnelheden (Figuur 5-4) zowel bij vloed als bij eb. Bij kentering zijn de stroomsnelheden en concentraties het laagst. Naarmate de stroomsnelheden stijgen, nemen de concentraties toe en wanneer de maximum stroomsnelheden zijn bereikt nemen de concentraties terug af. Al kan het zijn dat op een bepaald moment in een getijfase en locatie de concentraties afnemen bij een toenemende stroomsnelheid of toenemen bij een afnemende snelheid. Dit zal afhankelijk zijn van het meetpunt ten opzichte van de rivierbedding en dwarssectie.
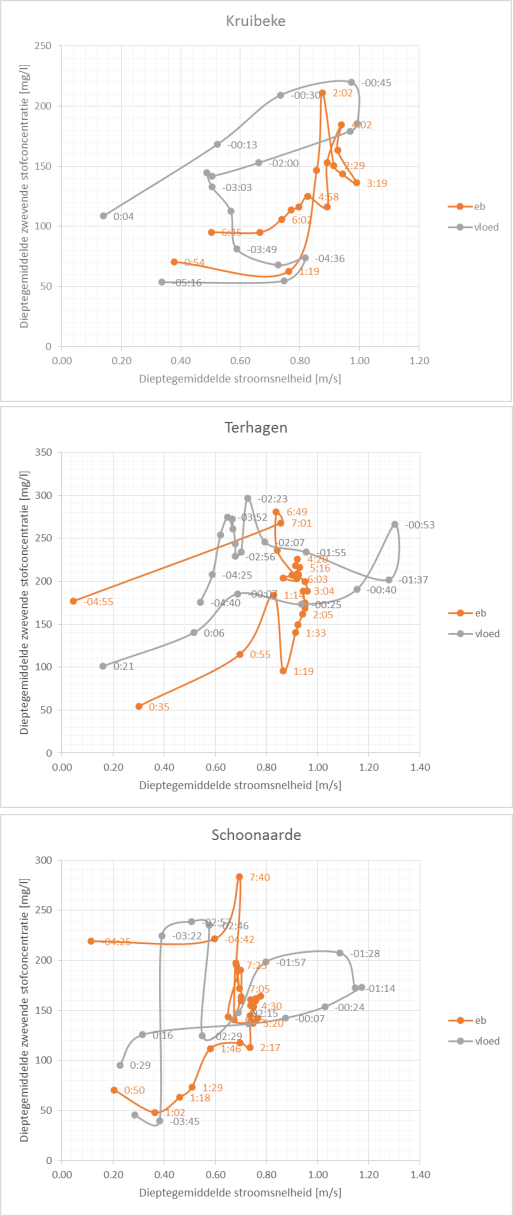
Figuur 5-4: Zwevende stofconcentraties in functie van de stroomsnelheden over een getij gemeten aan Kruibeke, Terhagen en Schoonaarde, met aanduiding van de tijdstippen voor hoogwater (uu:mm).
De waargenomen concentraties aan Schoonaarde zijn qua grootorde ca. 100 mg/l lager dan in 2014, maar de meetcampagne van 2014 was wel onder springtijcondities uitgevoerd. Anderzijds waren in Terhagen de waargenomen concentraties in 2015 hoger dan in 2014 (ca 50 mg/l). Er dient wel opgemerkt te worden dat de metingen van 2015 ter hoogte van Terhagen meer stroomopwaarts plaatsvonden zoals in 2012 en 2013. In 2011 en 2014 is meer stroomafwaarts gemeten wegens veiligheidsredenen. Bij vergelijking van de data met voorgaande campagnes dient hiermee rekening te worden gehouden. Op de andere meetlocaties is het verschil in positieligging kleiner, maar toch zijn er verschillen waarneembaar. Bijvoorbeeld zijn de snelheden lager aan meetpunten die meer aan de oever gelegen zijn. Dit is waar te nemen bij Kruibeke.
Figuur 5-5: Situeringkaart van de profielmetingen tijdens de tijcampagnes te Kruibeke, Terhagen en Schoonaarde.
5.2.2.GGG Lippenbroek
Tijdens een 13uurs meting te Lippenbroek, vond ook een monitoring van de zwevende stoffen aan Scheldezijde plaats tijdens het inwateringsmoment (4uursmeting) bij gemiddelde tijcondities. Deze resultaten zijn terug te vinden in Bijlage en worden hier verder besproken.
De profielmetingen op de Schelde tonen dieptegemiddelde stroomsnelheden tot 0.8 m/s. De maximale stroomsnelheid wordt 1 uur voor HW waargenomen en de minimale tijdens HW-kentering. De kentering vindt 0.5 uur na HW plaats en zet zich in aan de rechteroever (raaimetingen 1016 en 1018). Vanaf dit moment bezinkt het zwevende stof wegens de lagere stroomsnelheden naar de bodem (raai 1017) waardoor het concentratieverschil tussen wateroppervlak (42 mg/l) en bodem (260 mg/l) toeneemt. Ongeveer 1 uur na HW, wordt er terug meer zwevende stof opgenomen door de stijgende stroomsnelheden aan de rechteroever. Net voor dit moment, zijn de zwevende stofconcentraties in de waterkolom het laagst (lager dan 100 mg/l). De dieptegemiddelde concentraties aan het vast meetpunt variëren tussen 120 en 400 mg/l. De hoogste concentraties worden ongeveer 1 uur voor HW gemeten tijdens de hoogste stroomsnelheden.
De stroomsnelheden in de kil zijn lager dan de waargenomen snelheden in de Schelde. Doorgaans overschrijden snelheden niet 0.6 m/s en zijn er geen eenduidige richtingen waarneembaar. De gemeten concentraties zijn doorgaans lager dan op de Schelde en zijn homogeen verspreid. De hoogste concentraties zijn waarneembaar aan de ingang van de kil ongeveer 1 uur voor HW van ca. 300 mg/l. Tijdens de HW-kentering nemen de concentraties af tot waarden onder 100 mg/l.
Figuur 5-6: Situeringkaart van de profielmetingen tijdens de tijcampagnes te Lippenbroek
5.3.Lokale variatie over de dwarssectie
Binnen OMES is gekozen om zwevende stof te registreren op een vast meetpunt maar zwevende stof en stroomsnelheid varieert over de dwarssectie. Om de relatie van het vast meetpunt tot het de dwarssectie in kaart te brengen, worden er aan 9 stations dwarsraaien gevaren met een ADCP. De dwarsraaimetingen afwaarts gelegen van Antwerpen, tonen aan dat de waargenomen concentraties aan het vast meetpunt vaak hoger zijn dan de gemiddelde concentratie gemeten over de dwarsraai (Figuur 5-8). Dit verschil kan oplopen tot ca. 60%. Aan deze locaties is het estuarium het breedst en zal leiden tot meer variatie over de dwarsraai. Zo wordt bijvoorbeeld aan meetpunt Grens eerder hogere stroomsnelheden waargenomen tijdens eb meer aan de rechteroever en tijdens vloed meer aan de linkeroever. Dit is een gevolg van de ligging van het meetpunt (en dwarsraai) ten opzichte van de bestaande eb- en vloedstromen. Deze stromen worden gevormd door de riviermorfologie (vb. bocht van Bath). Dit verschil in stroomgedrag in combinatie met bathymetrie veroorzaakt een ruimtelijke variatie in zwevende stof over de dwarssectie dat varieert over het getij (Figuur 5-7). De variatie zorgt ervoor dat stroomafwaarts van Antwerpen, het zwevende stof jaargemiddelde van de profielen hoger zal zijn dan die van over de dwarssecties.
Op andere meetlocaties is de grootteorde aan zwevende stof gelijkaardig op het vast meetpunt als over de dwarsraai (Figuur 5-8). Al dient opgemerkt te worden dat aan station Lippenbroek de gemiddelde concentraties gemeten tijdens eb en over een profiel ca. 25% lager zijn dan gemeten over een raai.
De relatie tussen stroomsnelheden over een gemeten dwarsraai en profiel vertonen een grotere variatie dan zwevende stof (Figuur 5-9). Al is de stroomsnelheid gemeten over een profiel aan 5 stations (Grens, Liefkenshoek, Kruibeke, Temse en Lippenbroek) vaak hoger dan over een dwarsraai.
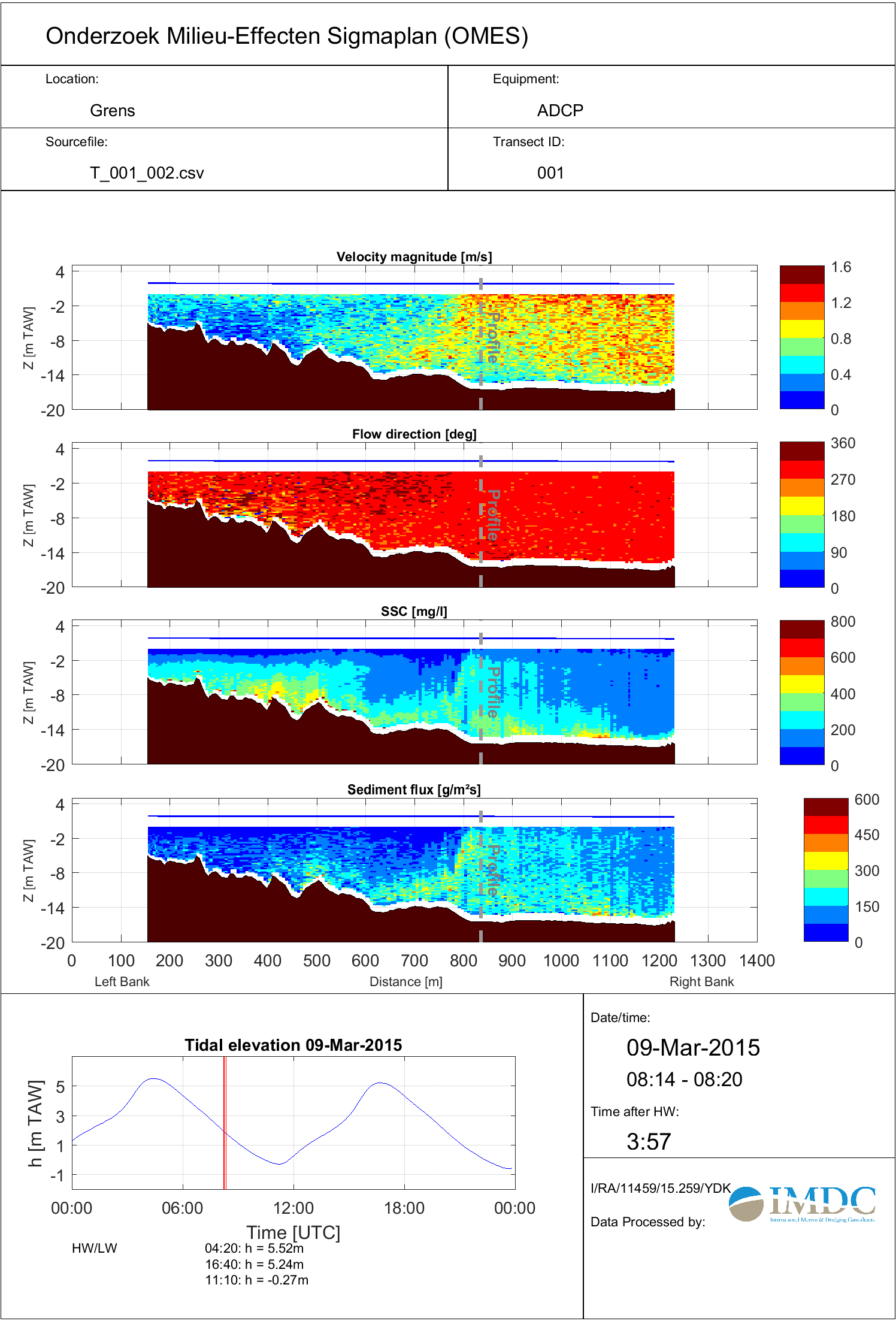
Figuur 5-7: Een voorbeeld van een dwarsraai aan station Grens op 09 maart 2015 tijdens eb.
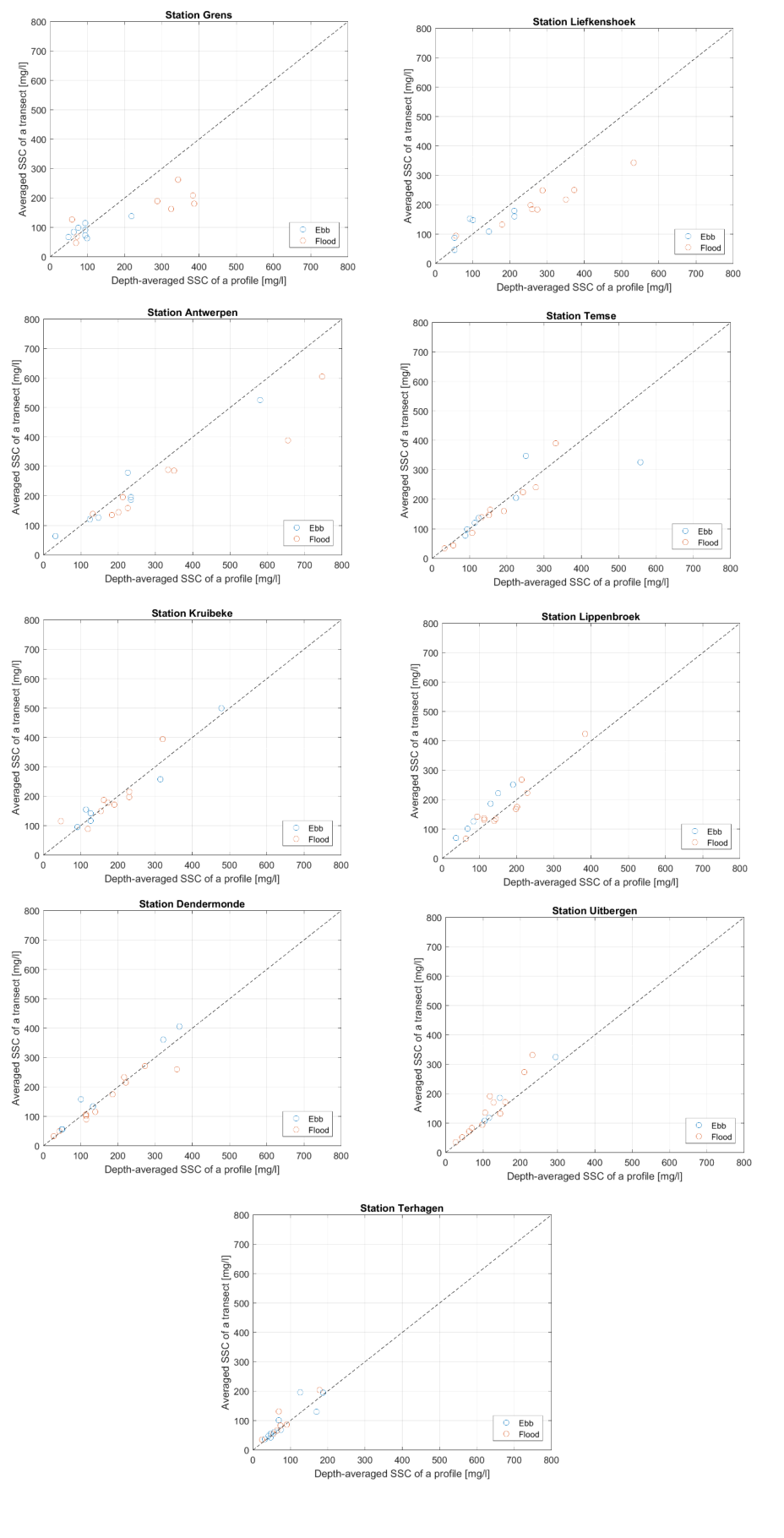
Figuur 5-8: Correlatie tussen zwevende stof langsheen een dwarsraai in functie van zwevende stof gemeten langsheen een verticaal profiel voor 9 OMES meetstations.
Figuur 5-9: Correlatie tussen stroomsnelheid langsheen een dwarsraai in functie van stroomsnelheid gemeten langsheen een verticaal profiel voor 9 OMES meetstations.
5.4.Verticale variatie
Niet alleen varieert de zwevende stofconcentratie over een dwarssectie maar ook over het verticaal profiel. Zo toont Figuur 5-10 de concentraties gemeten langs een verticaal profiel in Liefkenshoek (links) en Melle (rechts) als functie van de relatieve afstand boven de waterkolom. De profielen zijn weergegeven op basis van de ADCP-metingen (boven) en de dieptestalen (onder). In de grafiek worden alle metingen over de gehele duur van het meetprogramma (sinds 2002 voor de dieptestalen, sinds 2010 voor de ADCP-metingen) op deze meetlocaties onverwerkt weergegeven zonder uitmiddeling in de tijd of ruimte; er is bijgevolg een grote spreiding op de meetwaarden. Toch is op de grafieken een verticale gradiënt zichtbaar in de SSC-waarden, die sterker is in Liefkenshoek dan in Melle. De ADCP geeft bij elke meting en op elke locatie een volledig concentratieprofiel van ca. 20 datapunten, terwijl er bij staalnames slechts 2-5 stalen worden genomen op 1 locatie; derhalve zijn er in de ADCP-figuren meer datapunten dan in de figuren op basis van de staalnames.
Het Rouse-Vanoni-profiel is één van de meest courant gebruikte theoretische verticale profielen voor de sedimentconcentratie (Fredsøe en Deigaard, 1992):
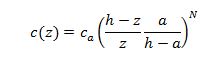
Hierin is
c de zwevende stofconcentratie
h de waterdiepte
z de afstand boven de waterbodem (positief opwaarts)
a een referentie-afstand boven de bodem (hier genomen als a= 0.01h
ca de referentieconcentratie op afstand z/a boven de bodem.
N is het Rouse-getal: ![]() , met ws de valsnelheid van het sediment, k de von Karman-constante en u* de schuifsnelheid (friction velocity).
, met ws de valsnelheid van het sediment, k de von Karman-constante en u* de schuifsnelheid (friction velocity).
Het Rouse-Vanoniprofiel is afgeleid voor uniforme, stationaire stromingscondities met een parabolisch viscositeitsprofiel; aannames die niet volledig geldig zijn voor de Zeeschelde. Toch geeft de formule een vrij goede fit met de data (Rouse-profiel in Figuur 5-10 valt bijna samen met de LOWESS-fit, een niet-parametrische regressie). Aangezien de aannames niet volledig geldig zijn moet het Rouse-getal N hier niet letterlijk worden gezien als ws/ku* , maar het kan nog steeds worden gebruikt om iets te zeggen over de vorm van het concentratieprofiel:
- N >> 1 wanneer hoofdzakelijk bodemtransport plaatsvindt (weinig suspensief transport)
- N =0 (1) duidt op suspensief transport met een duidelijke verticale gradiënt in het concentratieprofiel
- N << 1 duidt op suspensief transport waarbij het sediment quasi perfect is gemengd in de waterkolom.
Voor elke meetlocatie werd het Rouse-profiel gefit op de SSC-metingen. Figuur 5-11 (links) geeft het Rouse-getal N weer voor de verschillende meetlocaties in functie van de afstand tot de monding, op basis van de staalnames. Er is een duidelijke trend zichtbaar waarbij het Rouse-getal afneemt naarmate men verder opwaarts gaat. In het opwaartse deel van de Zeeschelde is N << 1 (sediment bijna uniform verdeeld over de waterkolom), in het afwaartse deel van de Zeeschelde is N~0.1 à 0.2: er is een verticale gradiënt in het concentratieprofiel maar deze is niet extreem sterk.
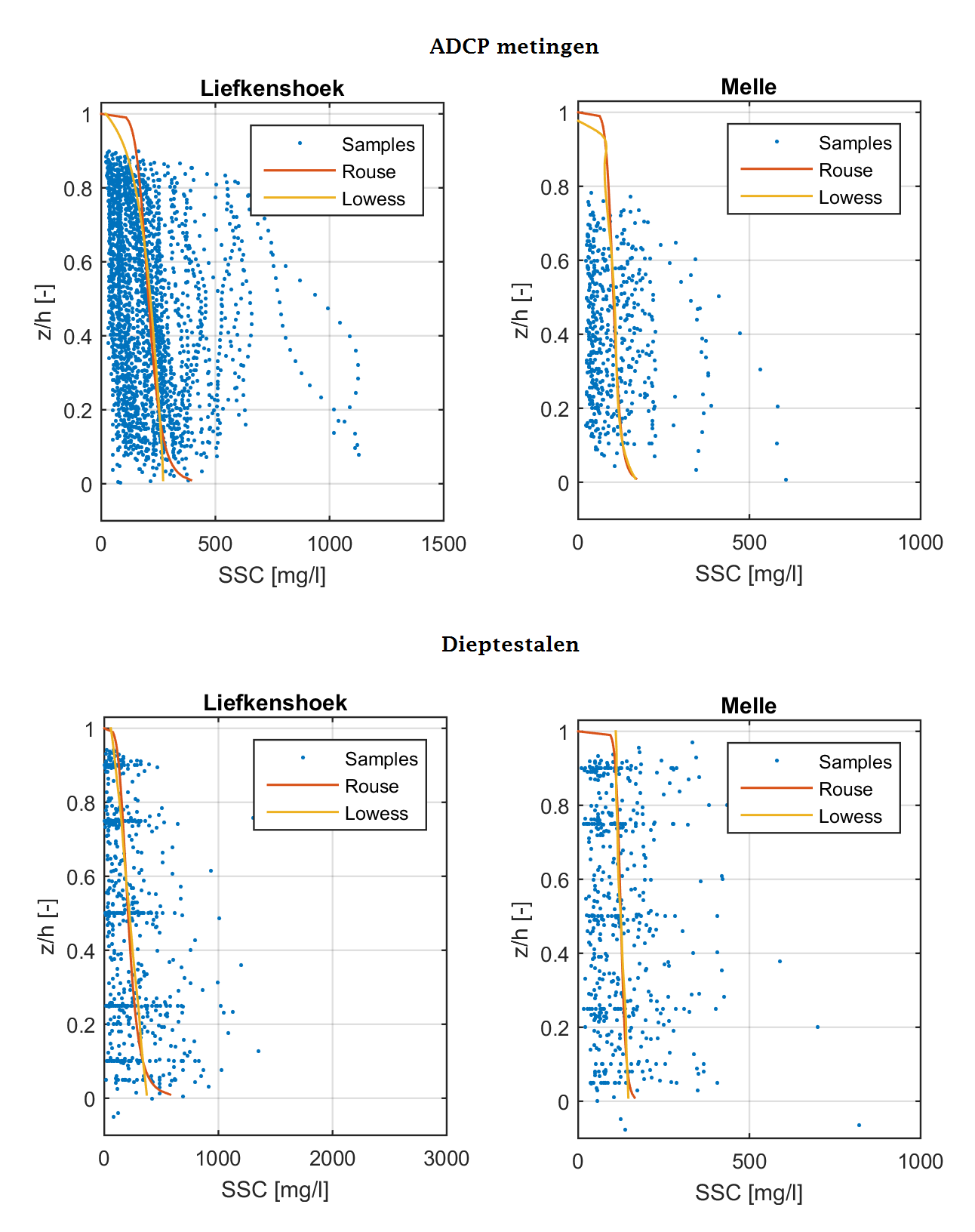
Figuur 5-10: Verticale verdeling van de SSC voor Liefkenshoek (links) en Melle( rechts). Profielen zijn weergegeven op basis van de dieptestalen (boven) en ADCP-metingen (onder).
Een andere manier om het verticale concentratieprofiel te beschrijven is de gemiddelde concentratiegradiënt (d(SSC))/dz . Deze parameter wordt bepaald door een rechte lijn te fitten door alle meetwaarden op een bepaald station en een bepaald jaar; de helling van de lijn is dan (d(SSC))/dz. De concentratiegradiënt is meestal negatief (SSC stijgt naarmate men lager in de waterkolom meet). Figuur 5-11 (rechts) geeft de gemiddelde concentratiegradiënt weer, genormaliseerd door de gemiddelde concentratie, in functie van de afstand tot de monding. De concentratiegradiënt is bijna 0 in het opwaartse deel (sediment bijna uniform verdeeld over de waterkolom) en is sterker negatief in het afwaartse deel.
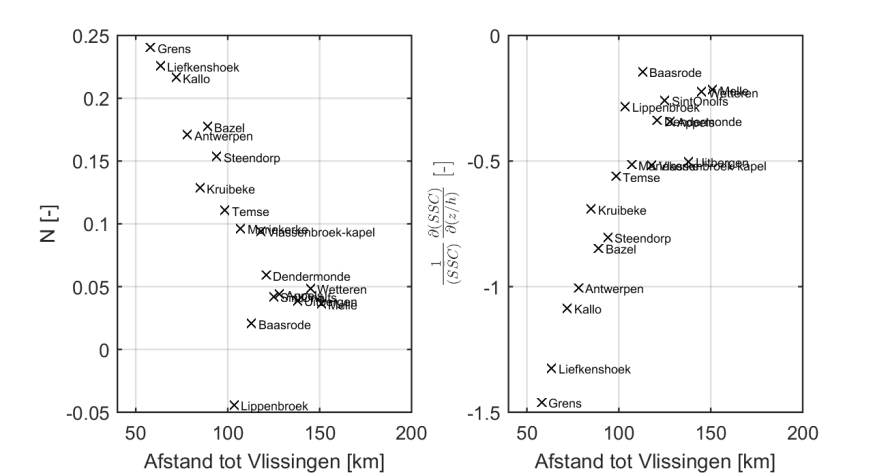
Figuur 5-11: Rouse-getal (links) en gemiddelde verticale concentratiegradiënt(rechts) in functie van de afstand tot de monding.
Het Rouse-getal en de gemiddelde verticale concentratiegradiënt zijn ook weergegeven in functie van de tijd in Figuur 5-12 (Beneden-Zeeschelde) en Figuur 5-13 (Boven-Zeeschelde). Telkens wordt bovenaan de evolutie op basis van de ADCP-metingen weergeven, en onderaan de evolutie op basis van de dieptestalen. De zwarte lijn geeft een LOWESS-fit weer door alle stations op basis van de ADCP-stalen. De magenta lijn geeft een LOWESS-fit weer door alle stations op basis van de dieptestalen.
In de Beneden-Zeeschelde (Figuur 5-12) vertoont de verticale concentratiegradiënt in de ADCP-metingen (grafiek bovenaan rechts) weinig evolutie doorheen de tijd, maar de ADCP-metingen beslaan enkel de periode 2011-2015. In de dieptestalen (grafiek onderaan rechts) neemt het Rouse-getal af en wordt de verticale concentratiegradiënt minder negatief doorheen de tijd; het sediment in suspensie wordt dus meer uniform verdeeld over de waterkolom over de jaren heen. Deze evolutie in de verticale concentratiegradiënt lijkt te zijn afgevlakt in de meest recente jaren 2011-2015. Om de resultaten op basis van de ADCP-metingen en de dieptestalen met elkaar te vergelijken, bevat de onderste rij figuren zowel de LOWESS-fit door de resultaten van de dieptestalen (magenta lijn) en de ADCP-metingen (zwarte lijn). Tijdens de periode waarin er voor beide datasets metingen zijn uitgevoerd komen de waarden uit de beide datasets goed overeen. De trend in de verticale menging wordt waarschijnlijk niet waargenomen in de ADCP-metingen omdat de tijdreeks te kort is.
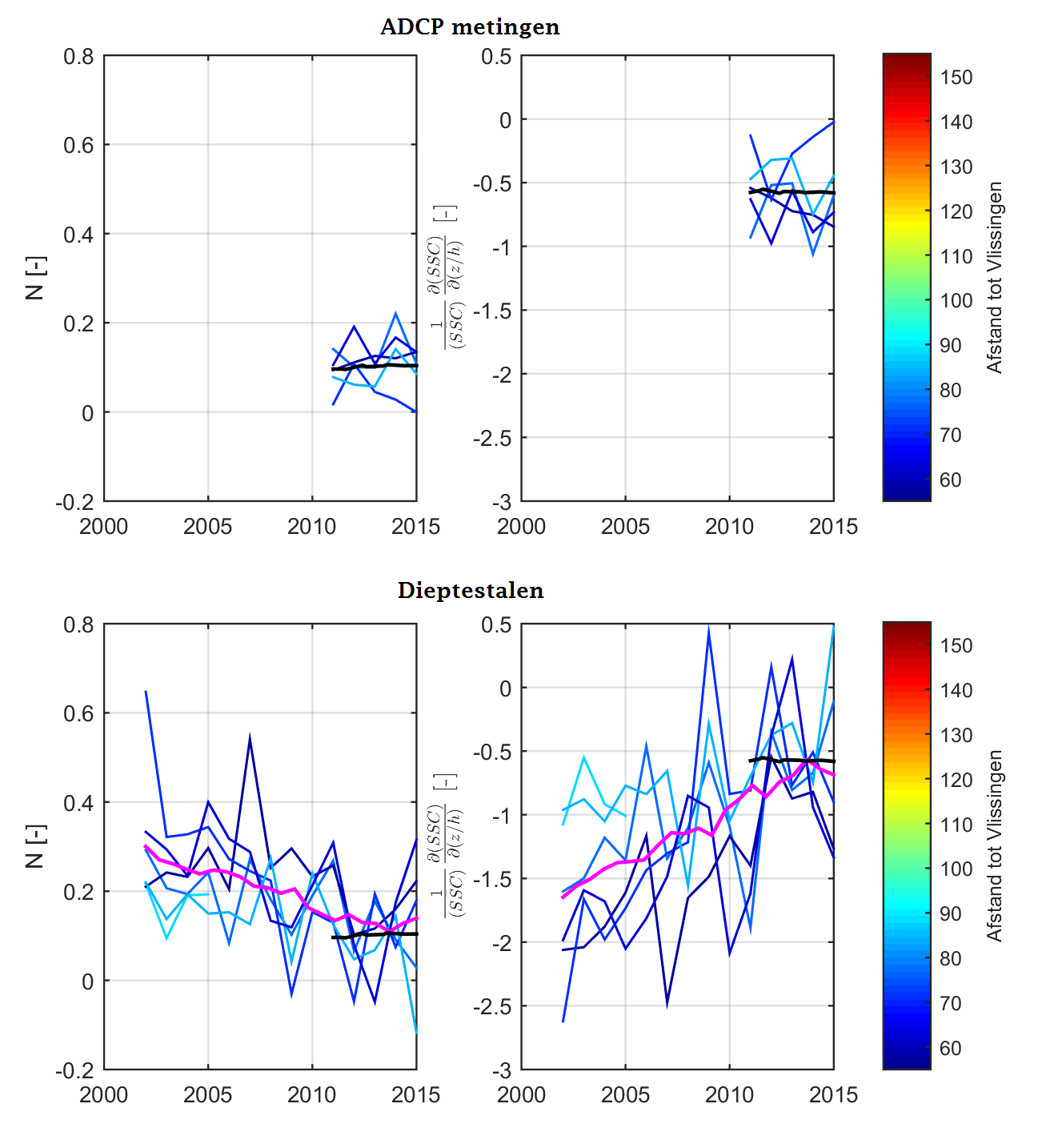
Figuur 5-12: Rouse-getal (links) en gemiddelde verticale concentratiegradiënt (rechts) in functie van de tijd voor de Beneden-Zeeschelde. Elke blauwe lijn stelt een meetstation weer, waarbij de kleurschakering de afstand tot Vlissingen weergeeft. De zwarte lijn is de LOWESS-fit door alle stations op basis van de ADCP-metingen en de magenta lijn is de LOWESS-fit door alle stations op basis van de staalnames.
In de Boven-Zeeschelde (Figuur 5-13) blijft het Rouse-getal en de verticale concentratiegradiënt eerder constant (misschien is er een lichte afname van het Rousegetal / toename van 1/((SSC) ) / (d(SSC))/dz). . Enkel in het jaar 2014 worden in de dieptestalen (onderste rij in Figuur 5-13) sterk afwijkende waarden waargenomen, waarbij de concentratiegradiënt zelfs positief wordt (SSC is hoger boven in de waterkolom dan onder in de waterkolom), en dit vooral in de meer opwaarts gelegen meetlocaties. In de meest recente jaren worden in de opwaartse stations echter minder dieptestalen genomen, en de afwijkende waarden in 2014 worden veroorzaakt door een relatief klein aantal datapunten die zijn gemeten tijdens dit jaar. In de ADCP-metingen (bovenste rij in Figuur 5-13) worden in 2014 geen afwijkende waarden waargenomen. Er wordt derhalve besloten dat er geen sterke evolutie in de verticale menging van sedimentconcentratie wordt waargenomen in de Boven-Zeeschelde.
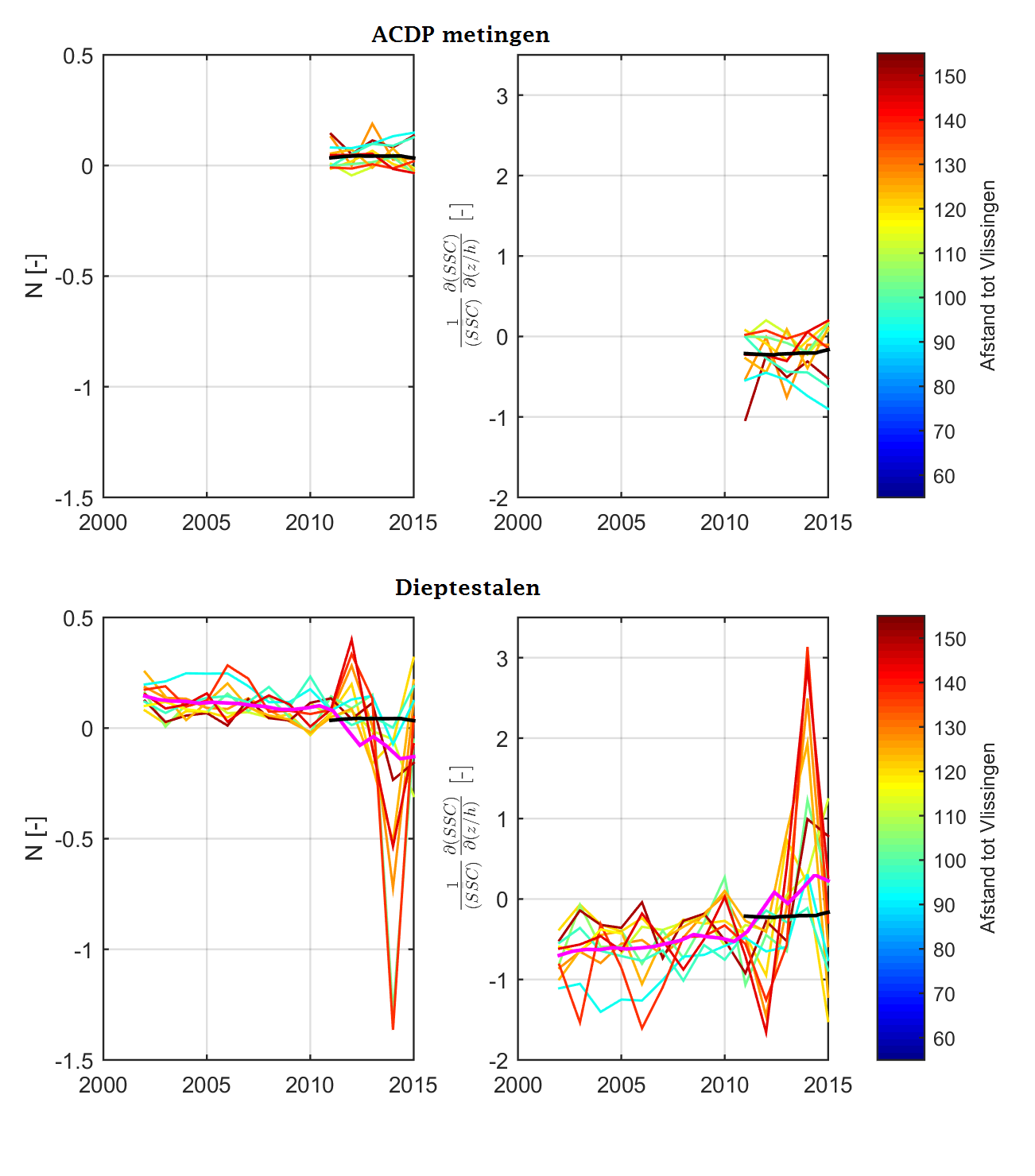
Figuur 5-13: Rouse-getal (links) en gemiddelde verticale concentratiegradiënt (rechts) in functie van de tijd voor de Boven-Zeeschelde. Elke lijn stelt een meetstation weer, waarbij de kleurschakering de afstand tot Vlissingen weergeeft. De zwarte lijn is de LOWESS-fit door alle stations op basis van de ADCP-metingen en de magenta lijn is de LOWESS-fit door alle stations op basis van de staalnames.
5.5.Lange termijn evolutie
5.5.1.Lichtextinctiecoëfficiënt kD en de ratio eufotische zone (Zeu) / mengdiepte (Zm)
Figuur 5-14 toont de eufotische diepte, afgeleid uit kD. Het lichtklimaat werd echter niet sinds de start van OMES opgevolgd. Pas vanaf 2002 werd gestart met de metingen van lichtklimaat, aanvankelijk enkel tijdens het groeiseizoen van de algen. In de wintermaanden werd niet gemeten. Vanaf 2010 zijn er jaarrond data beschikbaar. Op Figuur 5-14 zijn daarom alle datapunten aangeduid.
De zone met de geringste lichtdoordringing bevindt zich meestal rond km 100, waar tevens de tijamplitude een maximum kent. De zone met sterke saliniteitsgradiënt kende de grootste lichtdoordringing. Echter, de laatste 6 jaar is de eufotische diepte sterk afgenomen in deze zone. Figuur 5-15 toont de gemiddelde lichtextinctie in lente en zomer, voor de periode voor 2009 (2002-2006) en na 2009 (2009-2015). Vooral in de lente is er een sterke toename van de lichtextinctie in de gradiëntzone, zodat na 2009 deze zone de hoogste kD-waarden kent. In de zomer verschuift dit maximum opwaarts tot ca km 100. De lichtextinctie ligt ook hier hoger na 2009 dan voordien.

Figuur 5-14 Surface plot van de eufotische diepte. Elke zwarte stip is een meting.
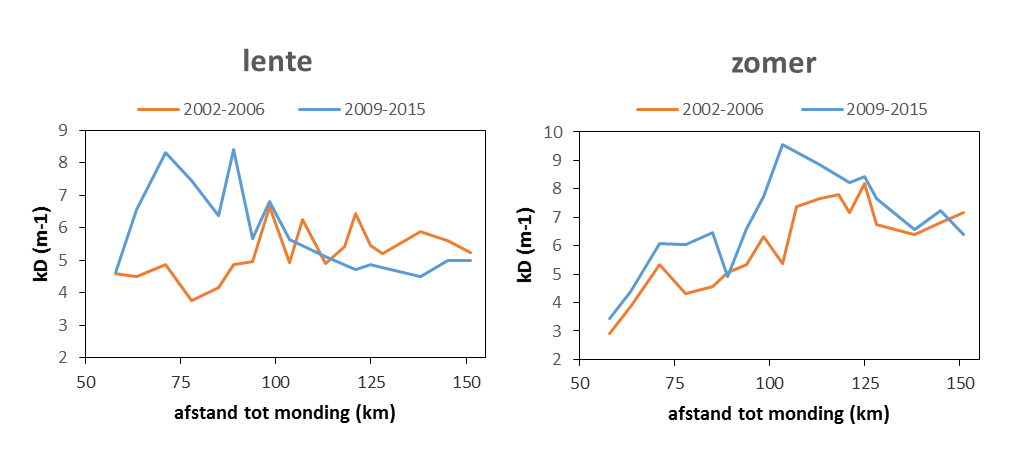
Figuur 5-15 Lichtextinctiecoëfficiënt kD (m-1) voor lente en zomer, in functie van de afstand tot de monding, voor de periode voor en na 2009.
In Figuur 5-16 zijn mediane en 90% percentielwaarden van Kd geplot. Er is flink wat variatie van jaar tot jaar, wat zou kunnen samenhangen met de fase van het getij waarin werd gemeten en met de debieten.
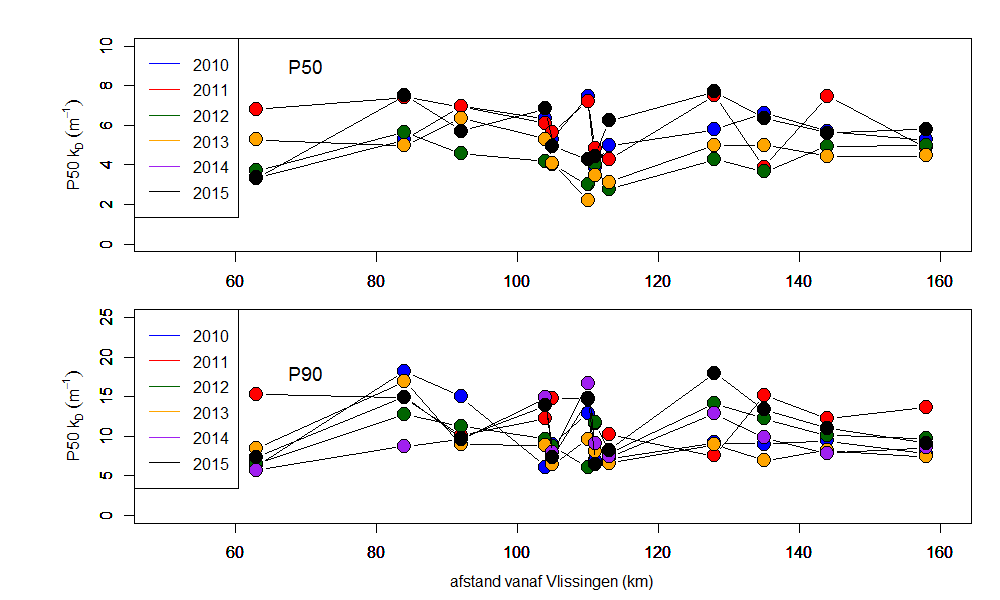
Figuur 5-16 P50 (boven) en P90-waarden voor de lichtextinctiecoëfficiënt Kd.
De laagste P50 waarden worden in het algemeen waargenomen in 2012 en 2013, dus dat lijken de jaren waarin het water relatief het helderst was. De P90-waarden zijn i.h.a. zeer hoog en tonen aan dat de fotische zone zeer beperkt kan zijn.
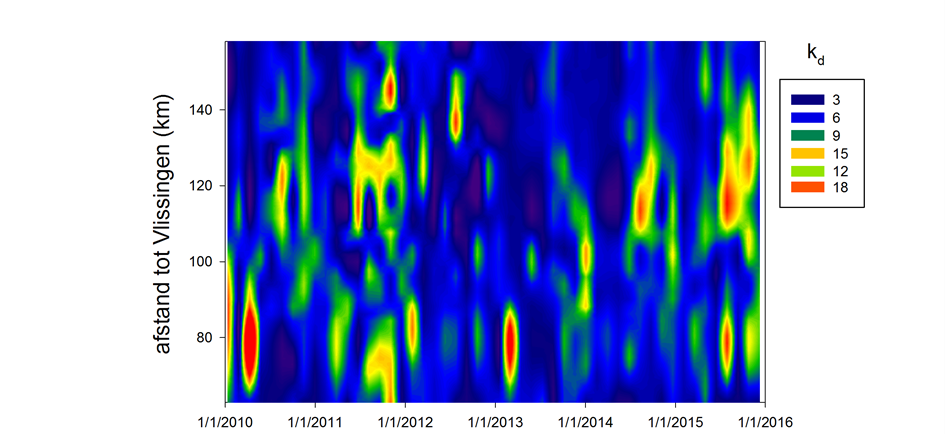
Figuur 5-17 contourplot voor Kd langsheen de Zeeschelde.
Uit Figuur 5-17 blijkt dat zeer hoge waarden over het gehele transect kunnen optreden en dit zowel in het voorjaar (2010, 2013) als in de zomer (2011, 2014 en 2015). Voor fytoplankton productie is echter niet de absolute Kd-waarde van belang maar de verhouding fotische diepte (Zeu): mengdiepte (Zm), waarbij Zeu berekend is als 4.6/Kd (waarbij Zeu de diepte is tot waar 1% van het licht doordringt). In Figuur 5-18 zijn de Zeu/Zm waarden weergegeven in een contourplot. Duidelijk is dat 2014 en 2015 een slecht lichtklimaat hadden, en alleen nabij Melle nog Zeu/Zm van groter dan 0.5 werden waargenomen.
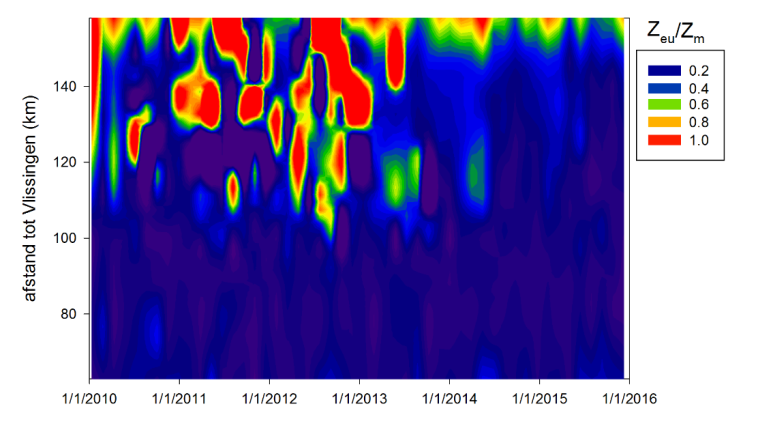
Figuur 5-18 contourplot voor Zeu/Zm langsheen de Zeeschelde
De mediane waarden verschillen in het algemeen weinig van jaar tot jaar maar nemen tussen Uitbergen en Melle toe (Figuur 5-19). De 90% percentielen geven veel meer spreiding: tussen de grens en Temse zijn de P90 vergelijkbaar met de P50 waarden en variëren rond de 0.2, wat betekent dat alleen in de bovenste 20 % van de waterkolom primaire productie mogelijk is. Tussen Dendermonde en Melle worden wel hoge P90 waarden gevonden en in 2010, 2012 en 2013 worden zelfs waarden van > 1 gevonden. Waarden > 1 betekenen dat de fotische diepte groter is dan de mengdiepte. In de Zeeschelde gaan we er van uit dat de ganse waterkolom goed gemengd is. Zeu kan in werkelijkheid dus niet groter worden dan Zm, waardoor Zeu/Zm een bovengrens kent van 1. Waarden waarbij de berekende Zeu > Zm impliceren dus dat er er licht tot op de bodem komt, ook tijdens hoog water. Dus ondanks de hoge troebelheid kan het lichtklimaat in het deel tussen Dendermonde en Melle in vooral 2011 en 2012 gunstig zijn omdat de gemiddelde diepte zo gering is. Zoals uit de contourplot van Zeu/Zm verwacht kon worden, zijn de P90-waarden ook voor het transect tussen Dendermonde en Uitbergen laag.
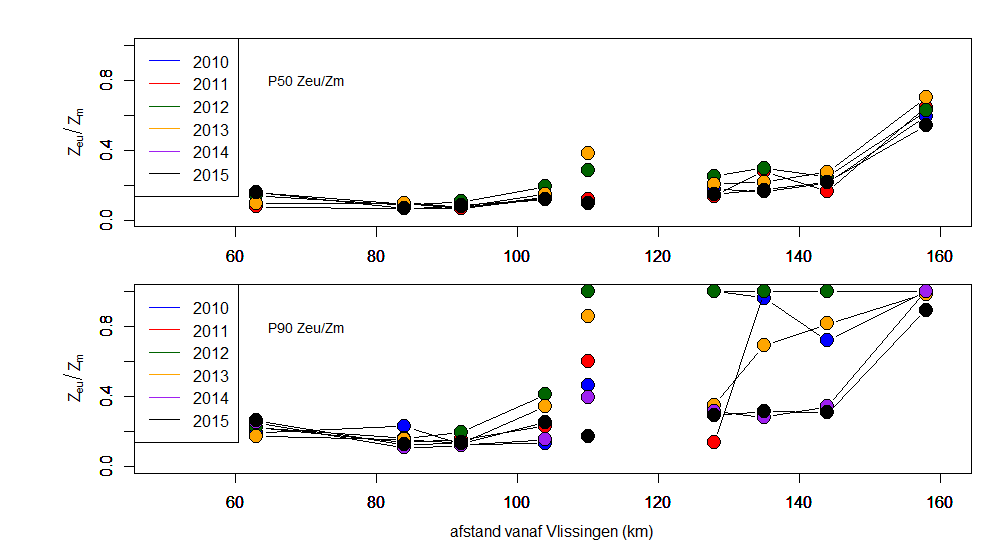
Figuur 5-19 P50 en P90 waarden voor Zeu/Zm. Let op de grote variatie in de P90 waarden in het deel bovenstrooms van Uitbergen (km 128) en Lippenbroek estuarien (km110). Voor de zijrivieren zijn geen data omdat daar de gemiddelde diepte onbekend is.
5.5.2.Zwevende stof nabij het oppervlak
Zwevende stof nabij het oppervlak wordt sinds de start van de OMES campagnes opgevolgd, door het nemen van schepstalen. De staalname en analyse werd in de loop der jaren door verschillende instanties uitgevoerd. Staalname gebeurde steeds op dezelfde, consistente wijze, maar de analysemethode toonde verschillen. Daarom werd een grondige data-analyse uitgevoerd op volgende databronnen:
- Lab. Analytische en Milieuchemie VUB – Baeyens (1996-2003)
- Lab. Ecologie en Systematiek, voormalige groep M.Tackx VUB (1996-1998)
- Lab. d'Océanographie Chimique et Géochimie des Eaux ULB, Vanderborght (2002-2009)
- Universiteit Antwerpen, Ecobe (2000-2015)
Uit de interkalibratie tussen de verschillende laboratoria blijkt dat de data van de 4 laboratoria goed tot zeer goed gecorreleerd zijn wanneer de specifieke geleidbaarheid onder 2000 µS/cm blijft (Figuur 5-20 en Figuur 5-21). Bij geleidbaarheden boven 2000 µS/cm zitten er echter significante afwijkingen op de spm resultaten. Voor 2005 is er vaak grote discrepantie tussen UA en ULB data. Oorzaak wordt gelegd bij onvoldoende naspoelen van de gefiltreerde stalen, waardoor zout op de filter achterblijft en werd meegewogen als ware het zwevende stof. Sinds 2005 is lab-procedure aangepast en is de correlatie wel goed. Ook bij data van het Laboratorium voor Analytische en Milieuchemie (VUB) en het Laboratorium voor Ecologie en Systematiek (VUB) zijn er gelijkaardige aanwijzingen dat er zout werd meegewogen bij de SPM analyse. Daarom werden alle SPM data bij SpCond >2000 van Ecobe, Lab. Analytische en Milieuchemie en Lab. Ecologie en Systematiek gemarkeerd als “suspicious” (verdacht) in de databank en niet verder meegenomen in de data-analyse.
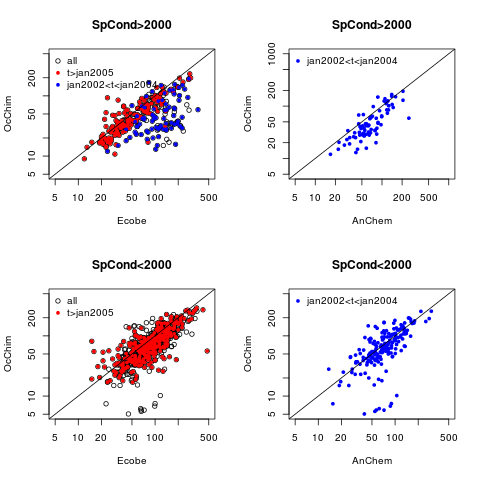
Figuur 5-20 Interkalibraties: Lab. d'Océanographie Chimique et Géochimie des Eaux ULB t.o.v. Ecobe (links) en t.o.v Lab. Analytische en Milieuchemie VUB (rechts) voor Specifieke Conductiviteit >2000 muS/cm (boven) en < >2000 muS/cm (onder). Merk: log-log plot
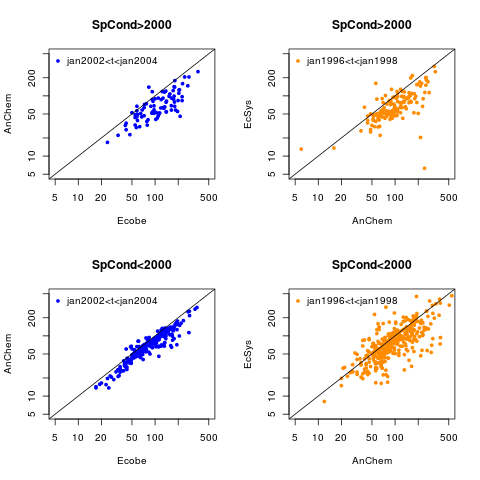
Figuur 5-21 Interkalibraties: Lab. Analytische en Milieuchemie VUB t.o.v Ecobe (links) en t.o.v. Lab. Ecologie en Systematiek (rechts) voor Specifieke Conductiviteit >2000 muS/cm (boven) en < >2000 muS/cm (onder) voor Specifieke Conductiviteit >2000 muS/cm (boven) en < >2000 muS/cm
5.5.3.Dieptestalen en ADCP-data
Bijlage presenteert de jaarresultaten van de waargenomen zwevende stof, stroomsnelheid en sedimentflux langsheen de 21 meetstations op de Schelde en de Rupel (Dijle en Beneden-Nete). Om een lange termijntrend te observeren, wordt de data uitgemiddeld tot jaargemiddelde (zie sectie 2.5.2) om de invloed van het getij op de concentraties te verminderen.
Met behulp van de ADCP kan op basis van de geregistreerde stroomrichting de tijfase tijdens de staalnames bepaald worden. In Tabel 5-1 wordt een overzicht gegeven van het aantal meetcampagnes per jaar en opgesplitst per getij. Sinds 2012 zijn er steeds 17 campagnes uitgevoerd over een meetjaar waarvan iets meer bemonsterd is tijdens eb dan tijdens vloed (uitgezonderd van meetjaar 2012). Deze asymmetrie kan een bias veroorzaken op de jaargemiddelden. Zo wordt vastgesteld in Figuur 5-22 dat de hoogste concentraties (> 400 mg/l) afwaarts van Antwerpen worden waargenomen tijdens eb. Op de andere meetstations wordt dit niet waargenomen, ook niet aan de meetstations op de Rupel waar stroomsnelheden hoger dan 0.8 m/s enkel worden waargenomen tijdens eb. Hieruit kunnen we afleiden dat de bias t.g.v. tijonafhankelijk bemonsteren op het jaargemiddelde vermoedelijk beperkt zal zijn voor meetstations opwaarts van Antwerpen.
Tabel 5-1: Overzicht van het aantal meetcampagnes over de tijd en opgesplitst per getijfase (eb, vloed en kentering) op basis van de ADCP-data.
| Totaal | Eb [min - max] | Vloed [min - max] | Kentering (<0.2m/s) | |
| 2011 | 13 | 5-11 | 2-7 | 0-2 |
| 2012 | 17 | 3-14 | 3-14 | 0-4 |
| 2013 | 17 | 7-11 | 4-7 | 0-2 |
| 2014 | 17 | 10-14 | 3-6 | 0-2 |
| 2015 | 17 | 8-13 | 4-9 | 0-5 |
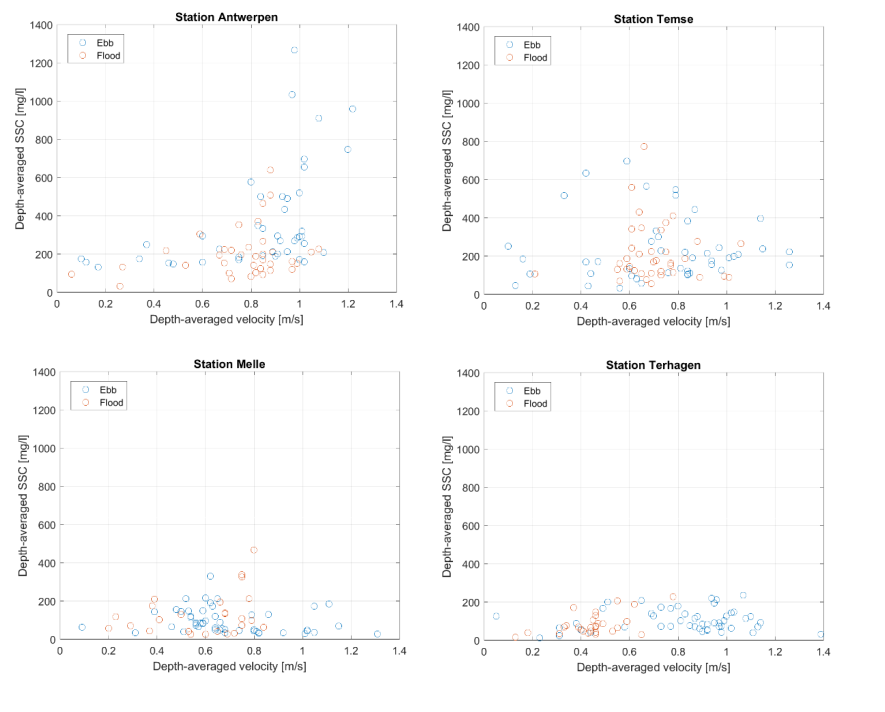
Figuur 5-22: Voorbeelden van dieptegemiddelde zwevende stofconcentraties in functie van de stroomsnelheid voor ADCP-data (2011-2015).
Een overzicht van de evolutie van de jaargemiddelden wordt weergegeven in Bijlage voor zowel ADCP als voor de dieptestalen. Vanaf 2011 worden de dieptestalen gebruikt om het akoestisch achtergrondsignaal van de ADCP om te rekenen naar zwevende stofconcentraties waardoor beide datasets een vergelijkbaar resultaat geven in evolutie. In absolute waarde, kan er een klein verschil optreden aangezien voor de ADCP een heel profiel wordt aanschouwd.
In de Boven-Zeeschelde varieert de jaargemiddelde concentratie tussen 60 en de 300 mg/l. De hoogste concentraties worden waargenomen in de omgeving van meetlocatie Steendorp en Temse. De jaargemiddelde concentraties zijn in 2004 en 2011 het hoogst en nemen daarna telkens af. In periode 2012-2014 zijn de concentraties zelfs de helft lager dan in 2011. Vanaf 2014 zijn de concentraties terug hoger maar lager dan in 2011.
In de Beneden-Zeeschelde (meetlocatie Kruibeke tot Grens) schommelen de jaargemiddelde concentraties tussen 100 en 400 mg/l. De concentraties zijn het hoogst in gebied van Liefkenshoek tot Antwerpen. De hoogste jaargemiddelde concentraties worden ook waargenomen in 2004 en 2011. In de periode 2005 – 2008 zijn de waargenomen concentraties het laagst. Na 2011 zijn concentraties gedaald tot in 2013 en zijn nadien terug toegenomen maar lager dan in 2011.
De jaargemiddelde concentraties op de Rupel variëren tussen +/- 50 en 200 mg/l en namen toe tot in het jaar 2011. Na 2011 nemen de concentraties af. De jaargemiddelde concentraties op de Dijle en Beneden-Nete zijn van gelijkaardige grootorde. De concentraties op de Zeeschelde nemen niet zichtbaar toe na de samenvloeiing van de Rupel (tussen meetlocaties Steendorp en Kruibeke) en de samenvloeiing heeft zichtbaar weinig effect op de zwevend stofgehalte op de Zeeschelde. De jaargemiddelde concentraties in 2015 in dit gebied zijn tussen 60 en 100 mg/l, wat gelijkaardig is aan de 2 voorgaande jaren. In 2014 waren er werkzaamheden ter hoogte van Niel aan de kaaimuur maar gezien de vergelijkbare resultaten mag aangenomen worden dat de invloed van de werkzaamheden beperkt waren.
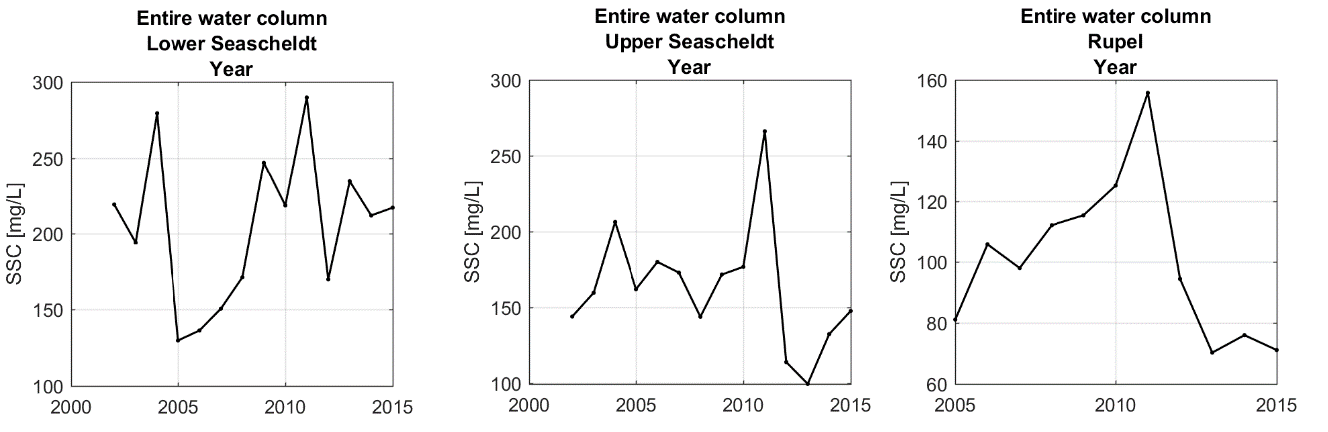
Figuur 5-23: De jaar- en systeemgemiddelde van zwevende stofconcentraties voor dieptestalen uit de volledige waterkolom voor Beneden-Zeeschelde (links), Boven-Zeeschelde (midden) en Rupel (rechts).
5.5.4.Troebelheid
Sinds meetjaar 2015 wordt er troebelheid waargenomen met behulp van optische backscatter-metingen (als deel van een multiparametertoestel). De meetresultaten worden gepresenteerd in bijlage. Gezien de korte meetperiode wordt deze niet besproken.
5.6.Evoluties in zwevende stof
5.6.1.Trends en trendbreuken
De zwevende stof nabij het oppervlak en in de diepte, op basis van de stalen, toont een zeer grillig verloop (Figuur 5-24). Dit kan deels verklaard worden door de tijonafhankelijk bemonstering. De surface plot met zwevende stof toont wel een aantal duidelijke trends. Ondanks de grote variatie, tekent zich duidelijk een maximum af in concentraties rond kilometer 100. Dit maximum neemt toe, en de zone met hoge concentratiewaarden wordt breder in opwaartse richting. Deze evolutie is analoog aan de trends in lichtextinctie. De zone met sterke saliniteitsgradiënt kende voor 2009 duidelijk lagere concentratiewaarden. Vanaf 2009 zijn de concentraties in ook deze zone sterk toegenomen. Dit wordt voornamelijk waargenomen in de oppervlaktestalen.
De dataset van de dieptestalen bevat echter slechts 7 meetjaren voor 2009 (een kortere baseline dan de dataset van de oppervlaktestalen, die teruggaat tot 1996), waardoor hier met minder zekerheid kan worden gesteld dat er zich een wijziging heeft voorgedaan in 2009. De (potentiële) wijziging na 2009 wordt voornamelijk waargenomen tijdens de winterperiode, en vooral in de bovenste 50% van de waterkolom. In Figuur 5-24 - Figuur 5-25 worden resultaten van oppervlakte- en dieptestalen met elkaar vergeleken.
De meetjaren 2009, 2011 en 2015 worden gekenmerkt door hoge concentraties. Dit zijn niet toevallig jaren met lage debieten. In de Beneden-Zeeschelde springen vooral de winters eruit, in de zoete zones worden maxima in de zomer waargenomen. Daarom zal de evolutie in concentraties voor en na 2009 beschouwd worden, in functie van seizoenen en debiet.
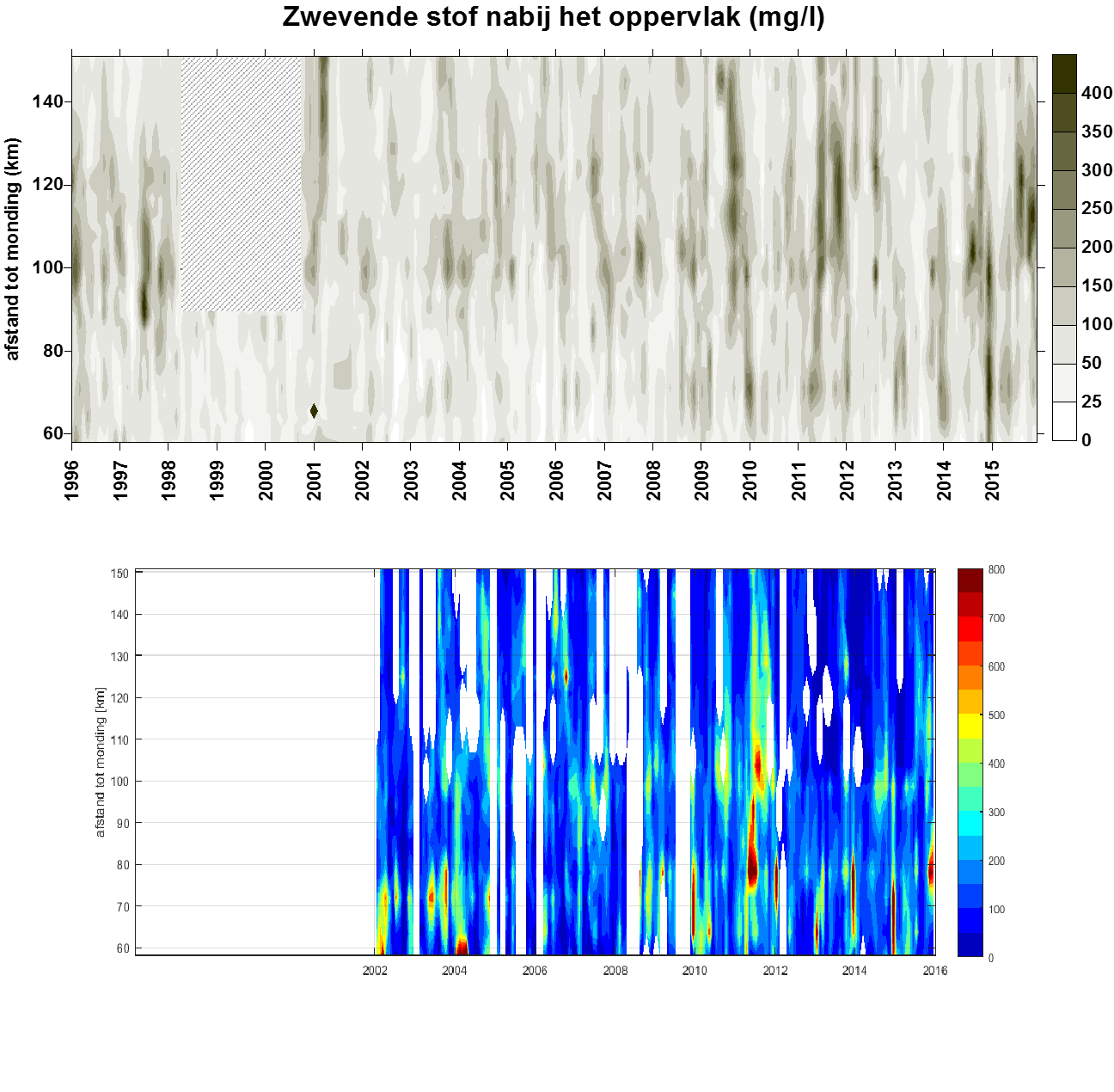
Figuur 5-24 Surface plot voor zwevende stof (mg/l) nabij het wateroppervlak (top) en dieptegemiddeld (onder), op basis van de stalen.
5.6.2.Invloed van seizoenale en bovenafvoer variaties
Voor 2009 lag het waargenomen maximum in zwevende stofconcentraties of turbiditeitsmaximum duidelijk in de zoete zone met lange verblijftijd, zowel in de winter als in de zomer (Figuur 5-25 en Figuur 5-26). Na 2009 zien we in het zomerhalfjaar een zeer sterke toename van zwevende stof, over de ganse Zeeschelde, maar vooral in de zoete zone met lange verblijftijd, waar de gemiddelde concentratie bijna verdubbelt aan het wateroppervlak. De toename van de concentratie sinds 2009 in de zoete zone is wel minder uitgesproken in de dieptestalen dan in de oppervlaktestalen. Dit kan mogelijk verklaard worden door het feit dat de tijdsreeks van de dieptestalen teruggaat tot 2002, en die van de oppervlaktestalen tot 1996 - er zijn dus minder meetjaren beschikbaar voor 2009. In het winterhalfjaar daarentegen, blijft de zwevende stofconcentraties in de zoete zone quasi gelijk, maar tekent zich een sterke stijging op in de gradiëntzone (km 60- 80).
De dieptestalen tonen aan dat de toename van de concentraties in de Beneden-Zeeschelde het sterkst is in de bovenste helft van de waterkolom, en minder sterk in de onderste helft van de waterkolom (Figuur 5-26). Dit hangt samen met de afname van de verticale concentratiegradiënt (SSC meer uniform over de waterkolom) die eerder werd aangehaald: de afname van de verticale concentratiegradiënt in de Beneden-Zeeschelde komt vooral doordat de SSC is toegenomen in de bovenste helft van de waterkolom.
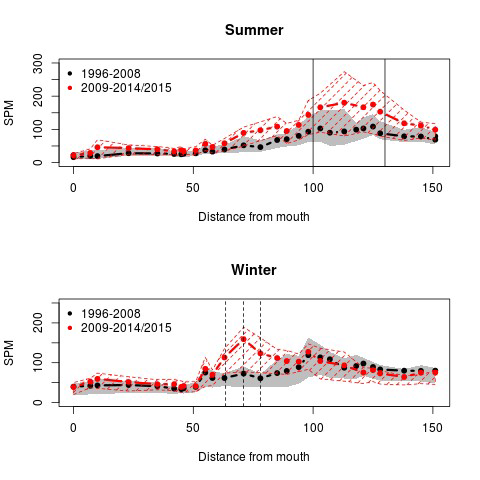
Figuur 5-25 Zomer- en winterhalfjaar gemiddelde concentratie aan zwevende stof (SPM, schepstalen) voor de OMES periode voor en na 2009, met aanduiding van de zone met turbiditeitsmaximum. De gearceerde en grijze band geven de spreiding op de data weer.
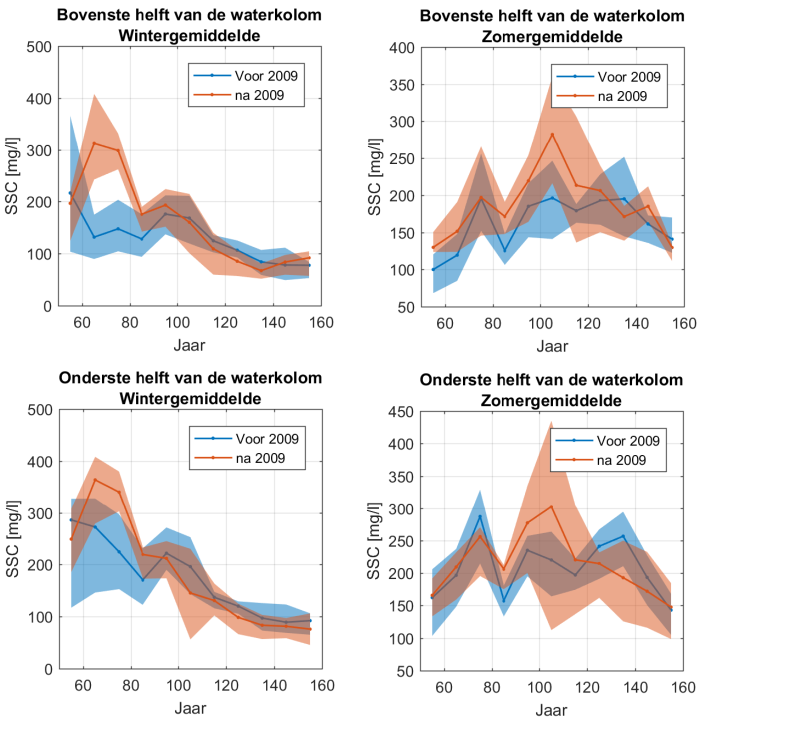
Figuur 5-26: Winter- (links) en zomergemiddelde (rechts) met aanduiding van 25ste en 75ste percentielen (gekleurde band) opgesplitst voor de bovenste (top) en onderste helft (onder) van de waterkolom.
Figuur 5-27 geeft de gemiddelde concentraties weer voor winter- en zomerhalfjaar voor de jaren in de periode voor en na 2009. De dunne lijnen stellen individuele jaren voor, de vette lijn het gemiddelde over de ganse periode. Zowel voor als na 2009, in zowel winter als zomer blijken er grote verschillen te zijn tussen de verschillende jaren onderling. Vooral in de zoete zone kunnen de verschillen tussen jaren zeer groot zijn, en zijn ze duidelijk gelinkt aan het netto zoetwaterdebiet. Bij lage debieten worden hogere zwevende stofconcentraties waargenomen. Een mogelijke verklaring hiervoor is dat het getij als het ware zwevende stof opwaarts opstuwt in het estuarium. Bij lage debieten kan het getij meer zwevende stof opstuwen in de zoete zone. Bij hoge debieten wordt de sedimentvracht meer afwaarts afgevoerd en dalen de concentraties in het opwaarts gedeelte.
Deze relatie tussen zwevende stof en debiet is echter grondig gewijzigd vanaf 2009: bij gelijkaardige debieten komen nu veel hogere concentraties voor. In de wintermaanden, bij hogere debieten, verschuift dit hogere turbiditeitsmaximum afwaarts en vormt vanaf 2009 in de gradiëntzone een wintermaximum.
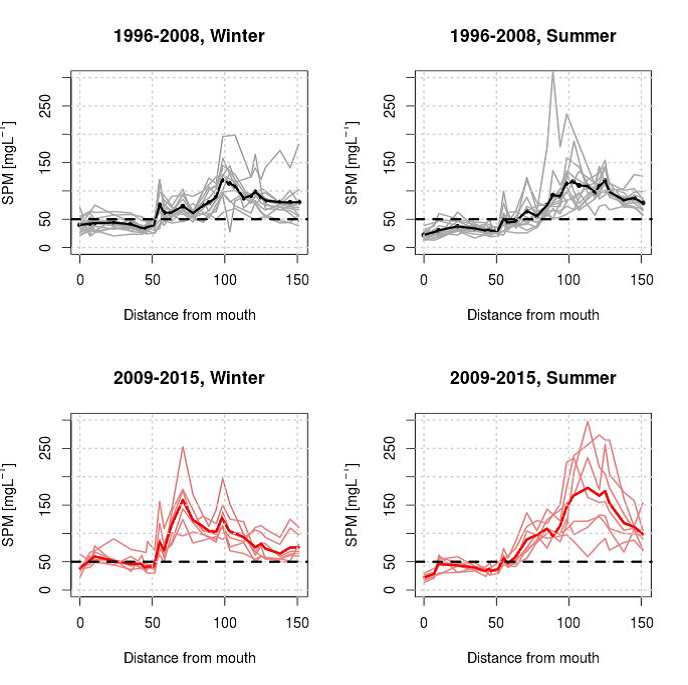
Figuur 5-27 Gemiddelde zwevende stofconcentratie voor winter- en zomerhalfjaar voor de jaren in de periode voor en na 2009. De dunnen lijnen stellen individuele jaren voor, de vette lijn het gemiddelde over de ganse periode.
De oppervlaktestalen in de zone tussen 100 en 130 km van de monding tonen voor 2009 een goede correlatie tussen zwevende stof en debiet in het zomerhalfjaar (Figuur 5-28). Na 2009 is deze correlatie grondig gewijzigd: de zwevende stofconcentratie in de zoete Zeeschelde is in de zomer ongeveer 8 maal gevoeliger voor debiet.
Figuur 5-29 geeft aan dat dit ook geldt voor de dieptestalen. De relatie lijkt gewijzigd voor zowel de zomer- als de winterperiode, en over de gehele waterkolom (zowel de onderste als de bovenste helft). De wijziging in de relatie is wel niet zo duidelijk als in de analyse van de oppervlaktestalen, omdat de dataset van de oppervlaktestalen over een lange periode loopt. De dataset van de oppervlaktestalen bevat meer datapunten in de periode vóór 2009, waaronder een aantal jaren met verhoogde bovenafvoer (in de dataset van de dieptestalen heeft enkel het jaar 2002 een verhoogde bovenafvoer). Daarnaast wordt opgemerkt dat de concentratie in het jaar 2009 waarschijnlijk hoger lag dan in de figuur wordt aangegeven, omdat er geen metingen zijn uitgevoerd tijdens een aantal maanden met hoge sedimentconcentratie.
De relatie tussen sedimentconcentratie en bovenafvoer werd ook onderzocht voor andere deelgebieden. Wanneer de volledige Boven-Zeeschelde wordt beschouwd, komt hetzelfde beeld naar voor als voor het deelgebied tussen 100 en 130 km van de monding. In de Beneden-Zeeschelde is er weinig of geen verband tussen bovenafvoer en sedimentconcentratie.

Figuur 5-28 Zomerhalfjaar gemiddelde zwevende stofconcentratie nabij het oppervlak (SPM) in de zoete zone (zoet korte en zoet lange verblijftijd) in functie van het zomerhalfjaar gemiddelde debiet te Melle (Q). In zwart zijn alle jaartallen voor 2009 weergegeven, in rood de jaartallen vanaf 2009.
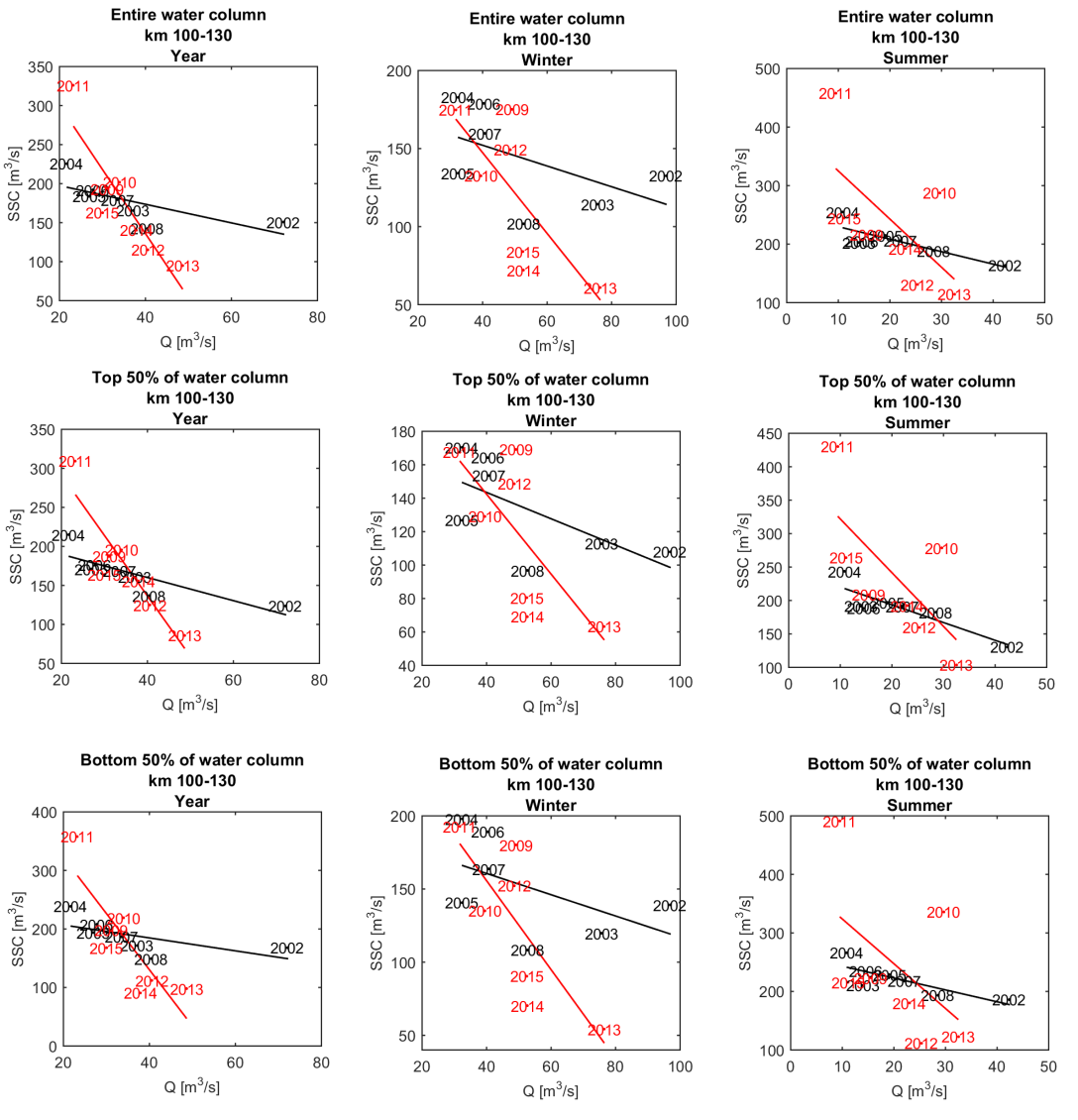
Figuur 5-29: Relatie tussen bovenafvoerdebiet en zwevende stofconcentratie (SSC) in het gebied tussen 100 en 130 km van de monding.
5.6.3.Invloed van baggeren en storten
In IMDC (2013, 2016) wordt aangetoond op basis van de continue meetreeksen aan meetstations (Boei84, Oosterweel en Driegoten) dat naast getij, bovenafvoer ook stortvolumes van slib afkomstig van de baggerwerkzaamheden hun invloed hebben op de zwevende stofconcentraties in de Zeeschelde. Deze relatie verloopt vermoedelijk in 2 richtingen. Bij toename in concentraties nabij sluistoegangen en dokken zullen de bagger- en stortvolumes ook toenemen en in andere zin zal bij toenemend stortvolumes de concentraties nabij de stortzones toenemen. Nabij de stortzones in de Beneden-Zeeschelde worden de concentraties beïnvloed op zowel korte als lange termijn ten gevolge van de stortvolumes. Zelfs aan het continue meetstation Driegoten (Boven-Zeeschelde) wordt een invloed op lange termijn waargenomen.
De relatie tussen jaar- of seizoensgemiddelde concentraties en stortvolumes van slib in de Beneden-Zeeschelde (stortzones Oosterweel, Plaat van Boomke, Punt van Melsele) werd daarom ook onderzocht op de OMES-datasets (Figuur 5-30). In de Beneden-Zeeschelde wordt er een relatie tussen de jaar- of seizoensgemiddelde concentraties en de stortvolumes van fijn sediment aan Plaat van Boomke, Punt van Melsele en Oosterweel. De relatie geldt zowel in de bovenste als de onderste helft van de waterkolom, maar is wel sterker in de onderste helft. De relatie geldt zowel tijdens de zomer- als de winterperiode.
Opvallend is wel dat er geen significante relatie wordt vastgesteld bij de stations afwaarts gelegen van de stortzones, uitgezonderd van station Liefkenshoek. Mogelijk wordt station Liefkenshoek beïnvloed door de baggerwerkzaamheden rondom Deurganckdok waarvan de baggerspecie hoofdzakelijk opwaarts wordt gestort en zorgt toch voor een (zwakke) relatie. Opwaarts van de stortzone voornamelijk bij station Antwerpen en Kruibeke, wordt wel een significante relatie gevonden tussen stortvolumes en zwevende stof. Bij hogere stortvolumes zijn de meetjaren aan zwevende stof ook hoger.
In de Boven-Zeeschelde is de relatie minder duidelijk. Indien meetjaar 2011 niet beschouwd wordt, wordt in de Boven-Zeeschelde eerder een omgekeerde relatie vastgesteld. Meetjaren met een lage zwevende stofgehalte in de Boven-Zeeschelde wordt gerelateerd met hoge stortvolumes in de Beneden-Zeeschelde. Of bij hoge debieten wordt de sedimentvracht meer afwaarts afgevoerd, dalen de concentraties in de Boven-Zeeschelde en bezinken deze eerder in de Beneden-Zeeschelde.
Er dient wel opgemerkt worden dat in meetjaar 2011 bijna 5 miljoen m³ aan materiaal werd gestort in de Beneden-Zeeschelde (Figuur 5-31). Het stortvolume in 2011 was voornamelijk hoger wegens aanpassing van de onderhoudsdiepte in Deurganckdok. Gecombineerd met een lage afvoer heeft dit vermoedelijk geleid tot een uitzonderlijk jaar aan zwevend stofconcentraties in de Zeeschelde (IMDC, 2016).
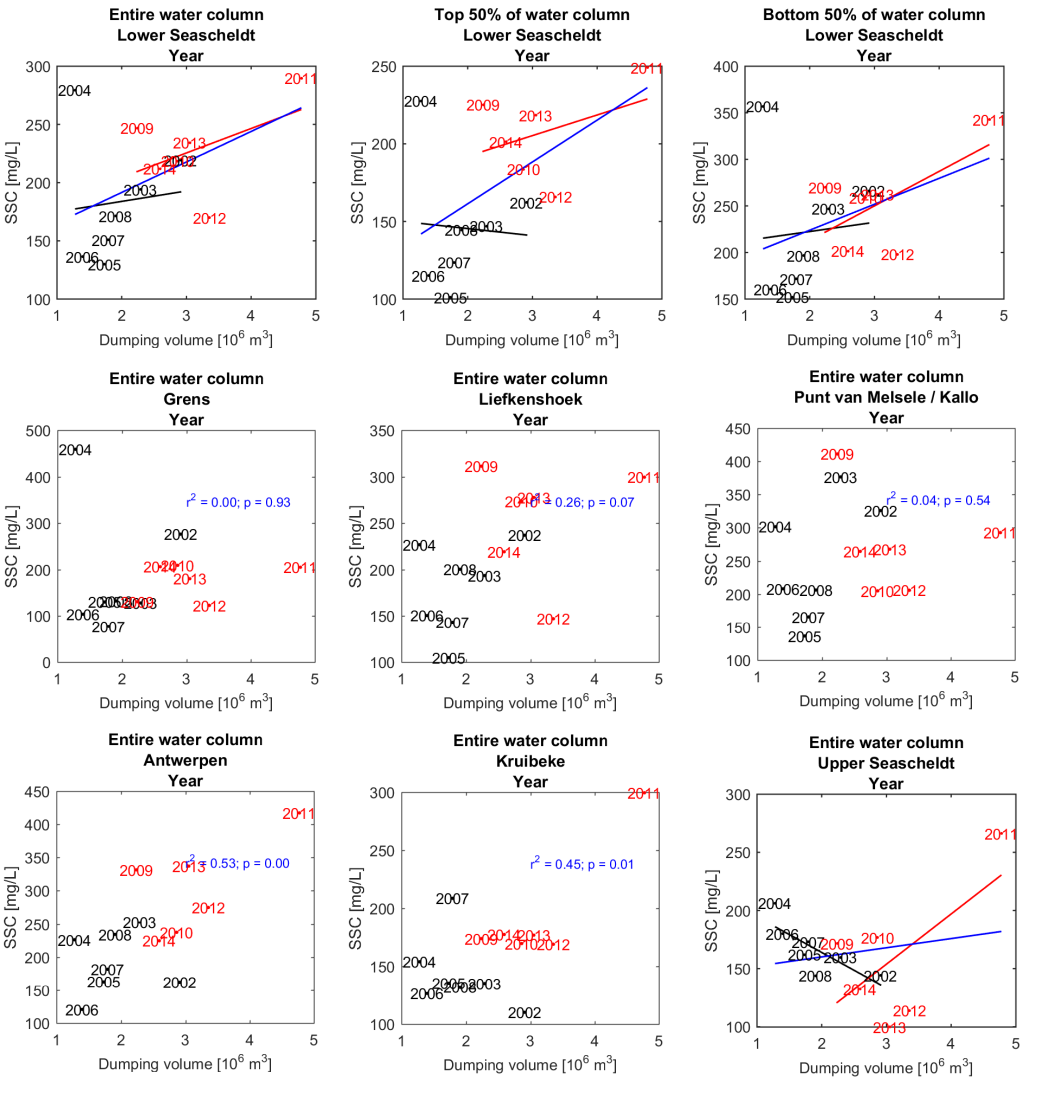
Figuur 5-30: Relatie tussen stortvolumes en zwevende stof in de Beneden- en Boven-Zeeschelde.
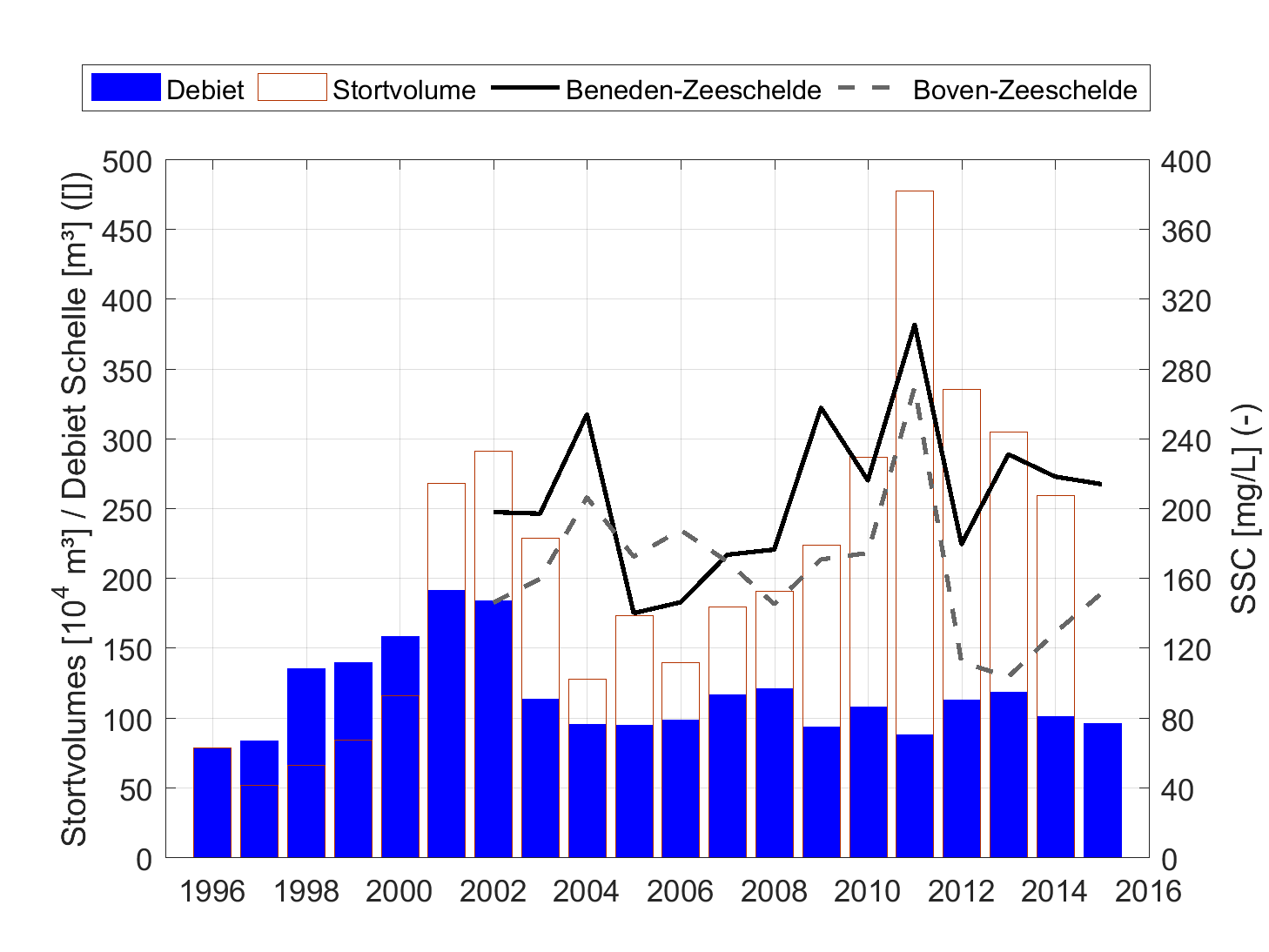
Figuur 5-31: Evolutie van het jaardebiet te Schelle, jaarstortvolumes in Beneden-Zeeschelde ( plaat van Boomke, punt van Melsele, Oosterweel) en van zwevende stof in Zeechelde op basis van dieptestalen.
Hoofdstuk 6.Plankton
6.1.Inleiding
Plankton wordt gedefinieerd als de verzameling van alle organismen die voornamelijk zwevend in het water leven, en zodoende voor hun (passieve) verplaatsing vooral afhankelijk zijn van de heersende stromingen. Binnen OMES worden zowel fyto- (plantaardig) als zoö- (dierlijk) plankton opgevolgd. Ook het fytobenthos, dit zijn de op de bodem of sediment levende algen, worden opgevolgd en mee in dit hoofdstuk gerapporteerd.
6.2.Fytoplankton: van een hypereutroof naar gewoon eutroof ecosysteem
6.2.1.Inleiding
De waterkwaliteit van de Zeeschelde is de voorbije jaren spectaculair verbeterd. In het hoofdstuk 4 Basis Waterkwaliteit werd deze verbetering toegeschreven aan de gestage verbetering van de kwaliteit in het bekken, ten gevolge van waterzuivering en reductie van stikstof en fosfaat, en het hierdoor herstelde zelfzuiverende vermogen van het estuarium. Dankzij een verminderde vuilvracht werd sterke algenbloei vanaf 2003 terug mogelijk, met sterke zuurstofproductie tot gevolg, waardoor in de Zeeschelde de resterende vuilvracht versneld verwerkt kan worden. Naast een evidente rol als basis van de voedselketen, speelt het fytoplankton bijgevolg ook een essentiële rol in het waterkwaliteitsverhaal van de Zeeschelde. Het Fytoplankton luik start daarom met de spectaculaire stijging van algenbloei in relatie tot de waterkwaliteit. In het volgende luik wordt vervolgens ingegaan op de evolutie van fytoplankton na de regime shift, waarbij de trends gedurende de laatste 6 jaar worden toegelicht.
6.2.2.Onderdrukking van de algenbloei
In de jaren ‘80 en ’90 was de zoete Zeeschelde zo sterk vervuild dat zelfs de groei van algen, het fytoplankton, werd geremd. Lage zuurstofconcentraties en hoge concentraties aan ammonium zorgden ervoor dat de zoete Zeeschelde niet alleen onleefbaar was voor de meeste hogere organismen (o.a. zoöplanktonsoorten, vis), maar dus ook sterke bloei van fytoplankton verhinderde. Algenbloei werd wel niet volledig uitgesloten, maar chlorofyl a, het belangrijkste pigment in algen, kende jaarrond veel lagere concentraties dan tegenwoordig (Figuur 6-1). Kortom, de Schelde was oververvuild of hypereutroof.

Figuur 6-1 Surface plot van de chlorofyl a concentratie (µg/l)
Literatuuronderzoek en modelberekeningen ondersteunen de hypothese dat algen belemmerd waren in hun groei door de slechte waterkwaliteit, hetzij rechtstreeks door de extreem lage zuurstofconcentraties, hetzij door andere kenmerken van de hypereutrofe toestand zoals hoge ammonium/ammoniak concentraties. Andere, klassieke verklarende factoren voor algenbiomassa, zoals licht- en nutriëntenbeschikbaarheid of uitspoeling verklaren de lange termijn trend in de chlorofyl a concentraties niet.
De nutriënten N, P en Si waren steeds in overmaat aanwezig (zie Waterkwaliteit). Door de slechte zuurstofomstandigheden was de zoöplanktonpopulatie niet optimaal (zie verder), zodat beperking van de algenbloei door begrazing weinig waarschijnlijk was. Algenbloei werd in de zoete Zeeschelde daarom vooral licht gelimiteerd verondersteld. Hoge concentraties aan zwevende stof (SPM) beperken de doordringing van zonlicht in de waterkolom (zie Lichtklimaat en zwevende stof), en bijgevolg de groei van algen. Echter, de sterke toename van chl a vanaf 2003 werd niet teruggevonden in een plotse afname van zwevende stof. Zowel voor als na 2003 kan er geen mooie correlatie opgetekend worden tussen SPM en chl a (Figuur 6-2). Ook uitspoeling van de algenpopulatie ten gevolge van hoge debieten biedt geen verklaring. De periode voor 2003 kent weliswaar een aantal zomers met extreem hoge debieten, maar ook deze blijken niet gecorreleerd met lage chlorofyl waarden.
Figuur 6-2 gemiddelde chl a concentratie (zomerhalfjaar) in de zoete zones in functie van debiet en zwevende stof.
Lage zuurstofwaarden kunnen wel algenbloei onderdrukt hebben: lage chl a waarden gingen samen met zeer lage zuurstofwaarden. Hoewel een direct negatief effect van anoxia op fytoplankton op het eerste gezicht tegenstrijdig lijkt (fytoplankton produceert immers zelf zuurstof), zijn er verschillende bekende mechanismen bekend van negatieve effecten van anoxia op het fotosynthetisch apparaat en op het groeimetabolisme. Herstel van fotoschade en foto-inhibitie verloopt trager, onvolledig of niet in anoxische omstandigheden. En zo goed als alle organismen hebben een zuurstofsensor die ervoor zorgt dat bij lage zuurstofconcentraties celfuncties worden vertraagd of stilgelegd. In de Schelde, die zeer goed gemengd is, vertoeven algencellen slechts een beperkt gedeelte van de tijd in de bovenste waterlagen waar licht kan doordringen (ongeveer ½ meter). De rest van de tijd vertoeven ze in de donkere, diepere waterlagen. In die omstandigheden produceren algen geen zuurstof en zal ook binnen de cel anoxia optreden wanneer dat in het omliggende water zo is, met de hoger vermelde negatieve effecten. Het netto-effect is een belemmerde groei van de algenpopulatie, iets wat ook experimenteel in het labo werd vastgesteld.
Een volledige verklaring biedt zuurstof echter niet. In de periode 1996-2002 komen algen immers wel enkele malen tot sterke bloei (chl a > 100 µg/l, Figuur 6-1), terwijl zuurstof in die periode steeds laag was. Een andere mogelijkheid is een onderdrukking van algenbloei door ammonium. Hoge concentraties NH4+ kunnen schadelijk zijn voor algen. Diatomeeën blijken bovendien gevoeliger aan NH4+ dan groenalgen (Collos & Harrison, 2014). Ammonium zelf kent een zekere toxiciteit, maar vooral de niet geïoniseerde vorm NH3 is schadelijk. Voor 2003 kwam deze stikstofvorm frequent voor in de zoete Zeeschelde (zie Waterkwaliteit), en speelde waarschijnlijk ook voor algenbloei een onderdrukkende rol.
Zelfs in lage concentraties zou ammonium een onderdrukkende rol kunnen spelen volgens de NH4+-hypothese die Dugdale en Wilkerson beschrijven voor San Fransisco Bay. Algen kunnen zowel nitraat als ammonium gebruiken als stikstofbron. Algen zouden een voorkeur hebben voor NH4+, maar het metabolisme, de groei zou trager zijn op deze stikstofbron. Als het aanwezige NH4+ uitgeput geraakt (< 0.004 mM of 0.06 mg/l), schakelen algen over op NO3 volgens de auteurs, waarna een snellere groei mogelijk is. Daarom werd ook in de Zeeschelde de invloed van ammonium op algenboei nagegaan.
In de zoete Zeeschelde komen de hoogste chl a waarden wel duidelijk voor bij lagere NH4+ waarden (Figuur 6-3). Deze surface plot geeft voor de zoete Zeeschelde chl a weer met een kleurschaal vanaf 100 µg/l om duidelijk de perioden van zeer sterke bloei te onderscheiden. De zwarte stippen zijn momenten waarop NH4 + zeer laag wordt (niet onder 0,06 mg/l maar onder 0,08 mg/l, zijnde de detectielimiet voor ammonium). Tijdstip en plaats waar de bloei start, vertoont overlap met lage ammoniumwaarden. In de Schelde zou dus NH4 + ook een rol kunnen spelen: pas wanneer NH4+ uitgeput geraakt, krijgen we maximale bloei.
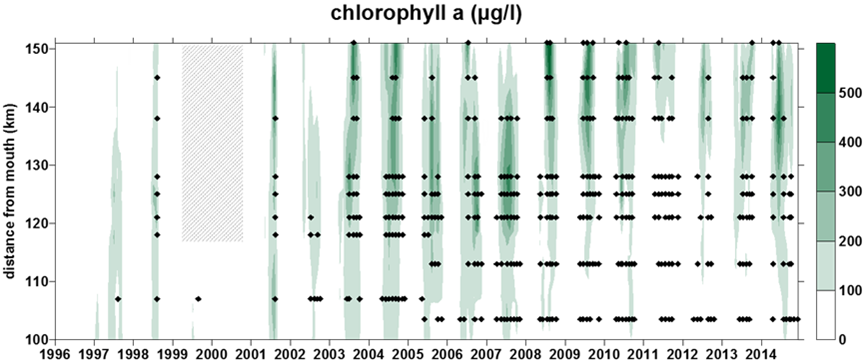
Figuur 6-3 Surface plot voor chlorofyl a. De zwarte stippen duiden op momenten waarbij de ammoniumconcentratie onder 0.08 mg/l duikt.
Echter, omdat bij sterke algenbloei veel zuurstof vrijkomt waardoor nitrificatie optimaal kan doorgaan, kan een lage NH4+ evenzeer een gevolg zijn van hoge chl a concentraties. Een vergelijking met de surface plot voor zuurstof (zie Figuur 4-1) toont echter dat hoge chl a of zeer lage NH4+ niet steeds even mooi samenvallen met hoge O2-waarden. Het samengaan van zeer hoge chl a en lage NH4+ is dus wellicht niet louter het effect van sterkere nitrificatie bij sterkere algenbloei (= zuurstofproductie).
Sterke algenbloei kan ook door opname van alle aanwezige NH4+ zorgen voor lage ammonium concentraties. Sterke algenbloei induceert dan lage NH4+ concentraties in plaats van omgekeerd. Om dit na te gaan, is gekeken naar de input van ammonium vanuit de Bovenschelde. De ammoniumconcentratie is de Zeeschelde (Melle) is immers afkomstig uit de Bovenschelde, maar algenbloei in de Zeeschelde kan geen invloed uitoefenen op NH4+ in de Bovenschelde. Bovendien zijn algenbloei (chl a) in Zeeschelde en Bovenschelde niet gecorreleerd (Figuur 6-4), zodat een sterke algenbloei met lage NH4+ in de Bovenschelde niet automatisch geïmporteerd wordt in de Zeeschelde.
Figuur 6-4 Scatterplot voor chlorofyl a (links) en ammonium (rechts) in Zeeschelde en Bovenschelde
Wanneer chl a in de Zeeschelde wordt uitgezet in functie van ammonium uit de Bovenschelde, blijkt dat meer hoge chl a waarden voorkomen bij de lage NH4+ concentraties (Figuur 6-5), maar een sterke aanwijzing voor de NH4+-hypothese vormt dit niet. Door echter te kijken naar de ammoniumvracht die vanuit Bovenschelde het estuarium binnenkomt, is het wel duidelijk dat enkel bij lage NH4+ -vrachten zeer hoge chl a waarden voorkomen (Figuur 6-5).
Echter, dit toont nog geen NH4+-hypothese aan: lage NH4+ -vrachten komen voor bij lage debieten. Een plot van chl a ten opzichte van debiet geeft hetzelfde beeld: er zal enkel sterke algenbloei zijn als het debiet voldoende laag is. Sterke algenbloei (>150 µg/l) komt te Melle enkel voor bij debieten onder 40 m³/s (Figuur 6-5).
Figuur 6-5 chlorofyl a in de Zeeschelde (Melle) in functie van de ammoniumconcentratie in de Bovenschelde, de ammoniumvracht vanuit de Bovenschelde en debiet.
Wanneer enkel deze lage debieten (< 40m³/s) worden beschouwd, blijkt een hoge chl a in de Zeeschelde onafhankelijk te zijn van debiet, maar wel van de ammoniumvracht. Sterke algenbloei (>150 µg/l) blijkt enkel voor te komen bij lage ammoniuminput (<40 g/s) (Figuur 6-6). Mogelijk inhibeerde dus ook in de Zeeschelde de aanwezigheid van ammonium een sterke algenbloei.
Figuur 6-6 Chlorofyl a in de Zeeschelde (Melle) bij lage debieten (< 40 m³/s) in functie van debiet (links) en in functie van de ammoniumvracht (rechts)
6.2.3.regimeshift
Het bijzondere aan het herstel van de zoete Zeeschelde is, dat de overgang van de oververvuilde, hypereutrofe toestand naar de huidige situatie relatief snel is gebeurd. Na decennia van weinig verandering, zagen we op enkele jaren tijd (ongeveer rond 2003) een omslag van een rivier met extreem lage zuurstofconcentraties in de zomer, naar een rivier met extreem hoge zomerse zuurstofcondities. Dat is een gevolg van de herstelde algenbloei. Voor 2003 was er door de grote zuurstofvraag te weinig O2 om alle NH4+ te nitrificeren en om sterke algenbloei toe te laten. Vanaf 2003 was de vuilvracht voldoende gedaald zodat algen opnieuw konden bloeien en bijgevolg ook extra zuurstof in het water konden pompen waardoor de vuilvracht versneld afnam. Het systeem sloeg om: de zogenaamde regimeshift (zie Waterkwaliteit). De zoete Zeeschelde is niet langer hypereutroof, maar vertoont nu de symptomen van een klassieke eutrofe toestand.
6.2.4.Bloei in het estuarium
Vanaf 2003 treed er jaarlijks sterke algenbloei op in de zoete Zeeschelde. In de nochtans zeer troebele Schelde komen de algen massaal tot bloei. Aangezien de concentraties in de Zeeschelde in de zomermaanden beduidend hoger liggen dan in de zijvieren, gaat het niet om louter import van algen uit het bekken, maar om algen die in het estuarium zelf tot sterke bloei komen (Figuur 6-7).
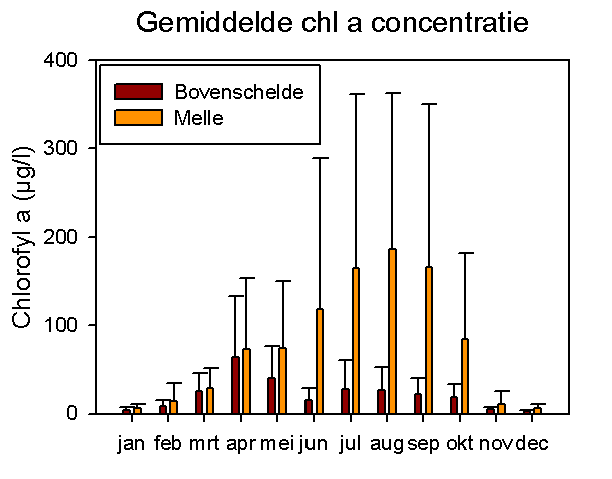
Figuur 6-7 Gemiddelde chl a concentratie in het estuarium te Melle en aan de boundary Bovenschelde
6.3.Fytoplanktonbloei in de eutrofe Schelde: analyse van de voorbije 6 jaar.
In de huidige Zeeschelde speelt de onderdrukking van algenbloei door de slechte waterkwaliteit (lage zuurstof, aanwezige ammonium) wellicht geen rol van betekenis meer. Hoewel er nog meer dan voldoende nutriënten zijn, treden er momenteel toch geen problemen op met overmatige algenbloei. Zo'n overmatige algenbloei kan de bekende negatieve effecten vertonen van eutrofiëring, zoals schommelingen in zuurstofconcentraties tussen dag en nacht (overdag produceren de algen zuurstof, 's nachts gebruiken ze daar zelf weer een deel van) en het mogelijk voorkomen van schadelijke algensoorten. Op dit moment veroorzaakt de algenbloei echter geen acute problemen. Wellicht vormt lichtklimaat nu steeds de limiterende factor. Bovendien kan de herstellende zoöplanktonpopulatie nu een rol van betekenis gaan spelen in het controleren van de algenbloei. Zoöplankton voedt zich immers met fytoplankton, en in vele gezonde estuaria is aangetoond dat de algenpopulatie door een combinatie van fysische randvoorwaarden en begrazing door zoöplankton wordt bepaald.
In de volgende paragrafen wordt de evolutie van fytoplankton besproken in de eutrofe, zuurstofrijke Zeeschelde gedurende de voorbije zes jaar.
6.3.1.Fytoplankton biomassa: chlorofyl a
Als maat voor de fytoplankton biomassa wordt de chlorofyl a (chl a) concentratie opgevolgd. De variatie gedurende het seizoen voor de verschillende jaren en stations is weergegeven als een serie boxplots in Figuur 6-8. Het chl a gehalte neemt toe vanaf station Grens en bereikt een maximum vanaf km 128 (Dendermonde), maar dit patroon varieert wel van jaar tot jaar: in 2010 en 2011 is er nog een toename in mediane waarden terwijl in de overige jaren de mediane waarden ongeveer gelijk blijven. Om de verschillen beter zichtbaar te maken zijn in de volgende figuren de 50 (P50 = mediaan) en de 90% percentielen (P90) geplot.
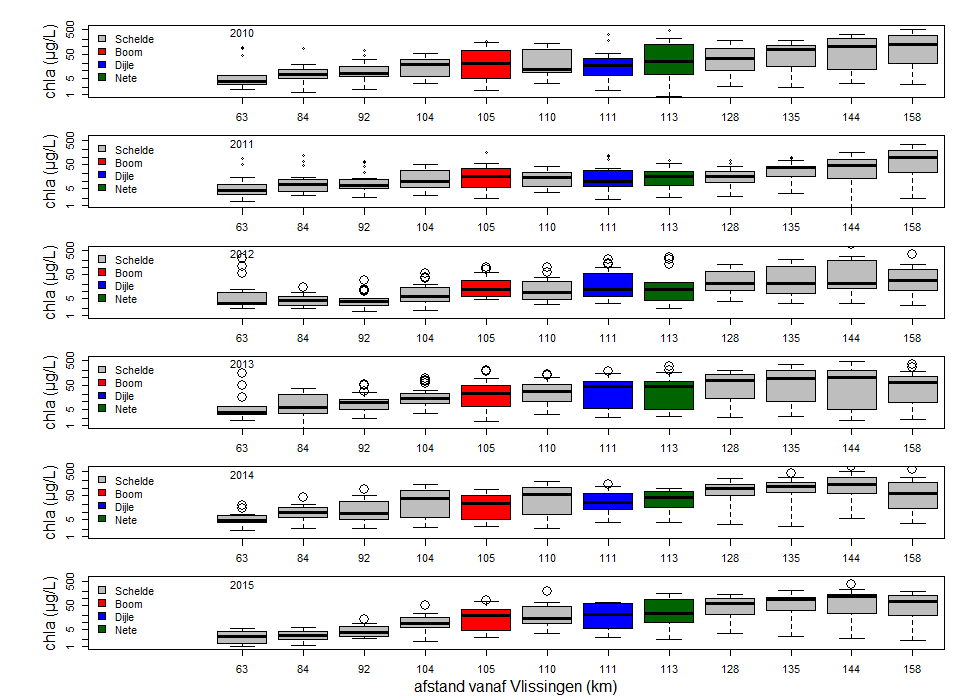
Figuur 6-8 Box-wisker plots van de variatie in het chla-gehalte als functie van de afstand tot Vlissingen. Stations in de zijrivieren zijn km 105 (Boom, in de Rupel), km 111 (Dijle) en km 113 (Nete) en zijn voorzien van een kleur.
De mediane (50% percentielen) waarden zijn geplot in Figuur 6-9. Duidelijk is dat 2012 het jaar is met de laagste mediane waarden, gevolgd door 2011. De hoogste mediane chl a concentraties worden gevonden in 2014. Opvallend is dat het er soms een toename is in chl a van Uitbergen naar Melle (km 144 tot km 158), terwijl in andere jaren er een sterke afname is.
In Figuur 6-10 zijn de 90% percentiel waarden (P90) van chl a geplot. Deze waarden geven een goede indruk van de intensiteit van de fytoplankton bloei. Er is geen duidelijk trend zichtbaar. Vanaf het Lippenbroek stroomopwaarts is 2011 het jaar met de laagste P90-waarden, terwijl dat jaar juist in Antwerpen en Kruibeke de hoogste P90 waarden worden genoteerd.
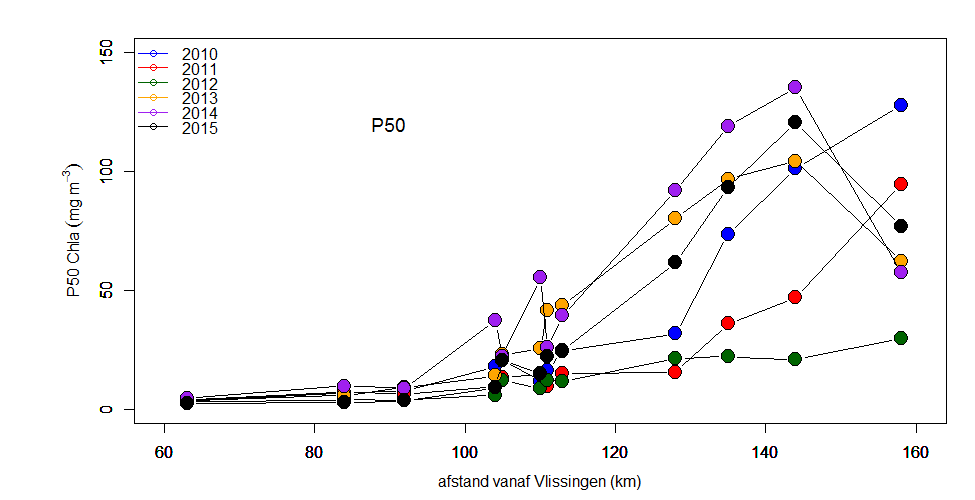
Figuur 6-9 Mediane waarden (P50) voor chla voor de jaren 2010 t/m 2015. Duidelijk is dat de laagste waarden werden gevonden in 2012 en de hoogste P50-waarden in 2014.
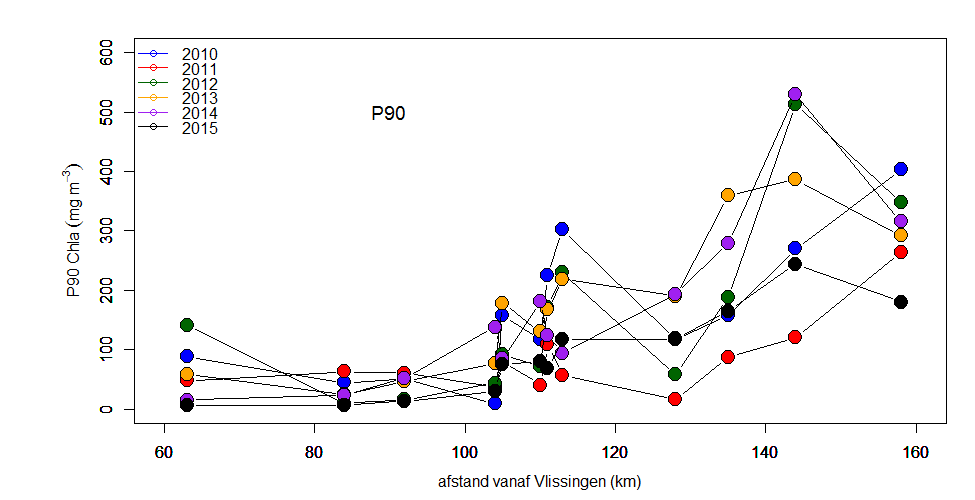
Figuur 6-10 P90 (90% percentiel) waarden voor chla.
Ter illustratie van het seizoensverloop zijn de chl a concentraties voor de stations Antwerpen en Melle geplot in Figuur 6-11. Voor Antwerpen geld dat de hoogste chl a concentraties werden waargenomen in 2010 en 2011, maar dat in die jaren de duur van de bloei zeer beperkt was tot enkele weken (2010) of een maand (2011).
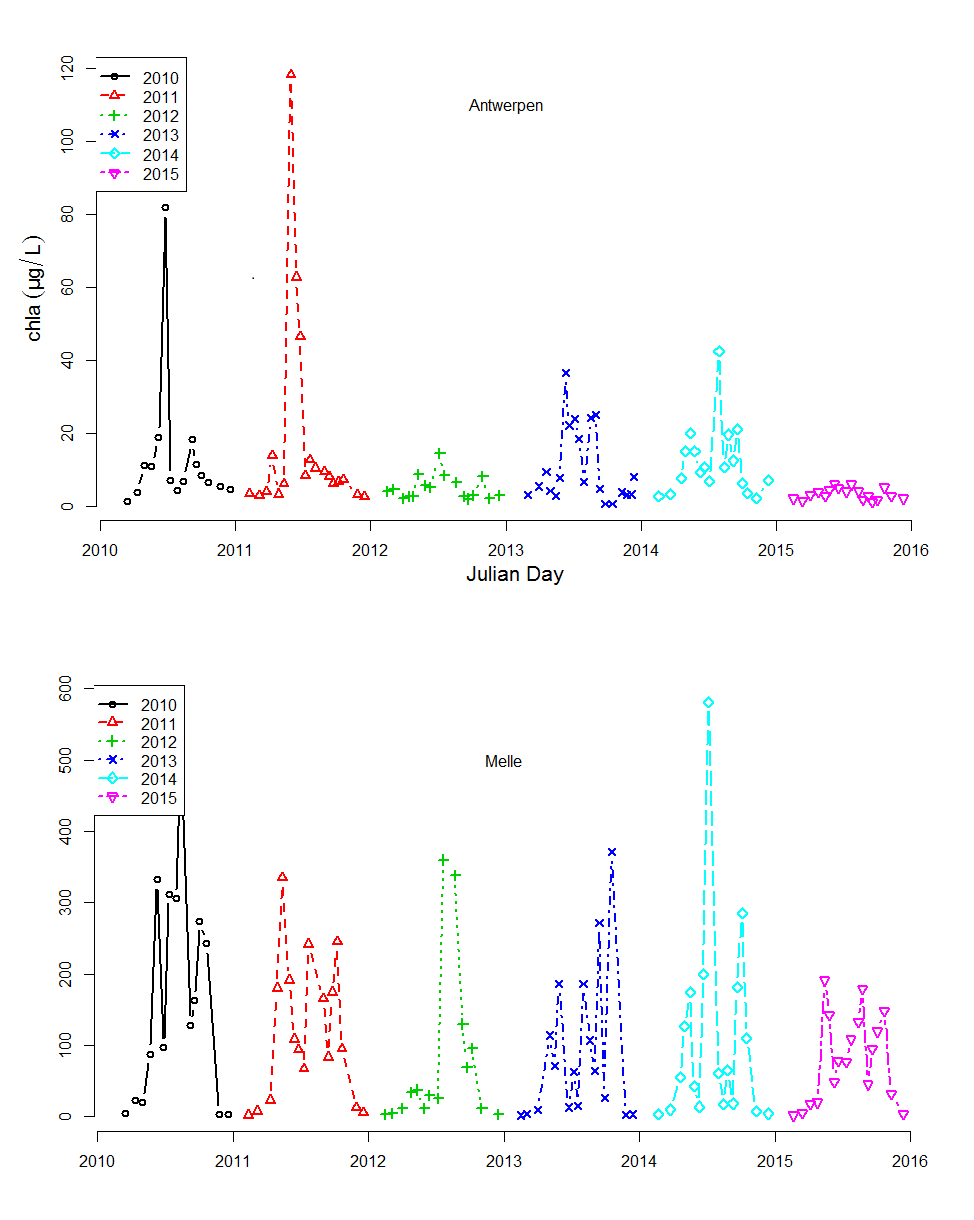
Figuur 6-11 Seizoensdynamiek in chla-concentraties voor de verschillende jaren voor de stations Uitbergen en Melle. NB: let op verschil in de waarde van de Y-as.
Deze grote verschillen tussen de verschillende jaren lijken veel minder voor station Melle en de bloei vertoont een grillig seizoenspatroon, maar de totale bloeiperiode lijkt duidelijk langer dan in Antwerpen. Voor de volledige periode zijn de chl a–concentraties geplot in een contourplot (Figuur 6-12). Vanaf ongeveer km 110 (station Lippenbroek) beginnen de hoge chl a concentraties en is de bloei ook duidelijk langer dan in het gebied tussen de grens en Rupelmonde. Opvallend is het ontbreken van een fytoplanktonbloei tussen de grens en Rupelmonde in 2015.
Figuur 6-12 Contourplot veranderingen in chl a in ruimte en tijd, de chl a concentratie (kleurschaal) is weergeven met log schaal.
6.3.2.Primaire productie: fotosynthese
Inleiding
Primaire productie is het proces waarbij algen, via fotosynthese, CO2 en nutriënten omzetten in nieuw celmateriaal. Zonlicht levert de hiervoor benodigde energie. De groei van algen wordt bepaald door bottom-up factoren als de beschikbaarheid van licht, nutriënten en de temperatuur. Tekorten aan macronutriënten in de Schelde zijn zeer onwaarschijnlijk: fosfaat en nitraat of ammonia zijn in overmaat aanwezig. Enkel opgelost silicium kan soms limiterend worden. Dit laatste nutriënt is essentieel voor diatomeeën, maar de andere groepen algen hebben hier geen behoefte aan. Colimitatie van verschillende nutriënten is mogelijk waarbij de ene soort/groep door een ander nutriënt beperkt kan worden dan een andere groep.
Omdat de Zeeschelde een troebel systeem is, zal licht meestal de beperkende factor zijn. Merk op dat limiterende factoren hierboven benaderd zijn als een fysiologische limitatie (Blackman type limitatie) en niet als een limitatie die de maximale “yield” bepaalt (Liebig’s law of the minimum), zoals in de landbouw gebruikelijk is. De reden hiervoor is dat de algenbiomassa zowel wordt bepaald door bottom-up factoren zoals hierboven beschreven, maar ook door top-down factoren als begrazing door schelpdieren en zoöplankton. Andere belangrijke verliesfactoren die de biomassa kunnen bepalen zijn sterfte (o.a. lysis door virusinfectie, een grote onbekende) en uitspoeling.
Omdat de turnovertime van het fytoplankton relatief kort is (enkele dagen) is de gemeten chlorofyl concentratie, de proxy voor biomassa, te beschouwen als de resultante van groei minus verliezen. Hierdoor is primaire productie een betere maat voor de maximale draagkracht dan de algenbiomassa (chlorofyl a) en worden binnen OMES metingen van primaire productie verricht. Hiertoe worden via lab experimenten de fotosyntheseparameters PBmax (maat voor de fotosynthese efficiëntie) en αB (maximale fotosynthese snelheid) bepaald (zie Hoofdstuk 2 voor de gebruikte methodologie), waarmee de primaire productie berekend kan worden.
PBmax
PBmax ( eenheid mgC (mg chla )-1 h-1) is de maximum fotosynthese snelheid per mg chlorofyl a. Figuur 6-13 geeft in een boxplot een overzicht van de variabiliteit in PBmax. In het algemeen lijkt de mediane PBmax het laagst in de zijrivieren, maar lage waarden worden ook waargenomen bij de grens, vooral in 2014 en 2015. Opvallend zijn de grote waarden tussen de P25 en P75 ("de grootte van de box") waarden gevonden voor Lippenbroek (km 110) in 2015 en voor Dendermonde (km 128) en Appels (km 135) in 2011 en 2012.
Figuur 6-13 boxplot van PBmax waarden voor de verschillende stations en de verschillende jaren.
Om het seizoenspatroon wat beter zichtbaar te maken, is het verloop in PBmax gedurende het seizoen weergegeven voor 5 stations in Figuur 6-14. Voor alle stations geldt dat PBmax een duidelijk seizoenspatroon volgt en de temperatuur lijkt te volgen. Alle stations hebben wel een paar hogere waarden > 15 mg C (mg chla)-1h-1 die niet in het patroon passen, zoals bij Zandvliet in 2016, in Antwerpen in 2014, in Dendermonde in 2011 of in Melle in 2013. Deze uitschieters zouden veroorzaakt kunnen worden door te lage chlorofyl concentraties, maar ze hebben geen invloed op de primaire productie omdat uiteindelijk de chl a concentraties geen rol spelen in de uiteindelijke uitkomst.
De PBmax waarden zijn ook weergegeven in een contourplot (Figuur 6-15). Uit deze contourplot blijkt duidelijk dat PBmax > 10-15 zelden voorkomen en zoals hierboven gemeld het gevolg lijken te zijn van uitschieters. De contourplot lijkt ook te laten zien dat er vrijwel geen ruimtelijke gradiënt is in de data. Om dit beter zichtbaar te maken, zijn de mediane waarden geplot in Figuur 6-16. Deze grafiek laat zien dat er toch een trend aanwezig is in de data, met de hoogste mediane PBmax waarden nabij de grens om tussen Antwerpen en Lippenbroek minimale waarden te laten zien. In het algemeen zijn de ruimtelijke patronen in PBmax echter beperkt.
Figuur 6-14 Timeplot of PBmax als functie van de dag voor de verschillende jaren voor 5 stations.
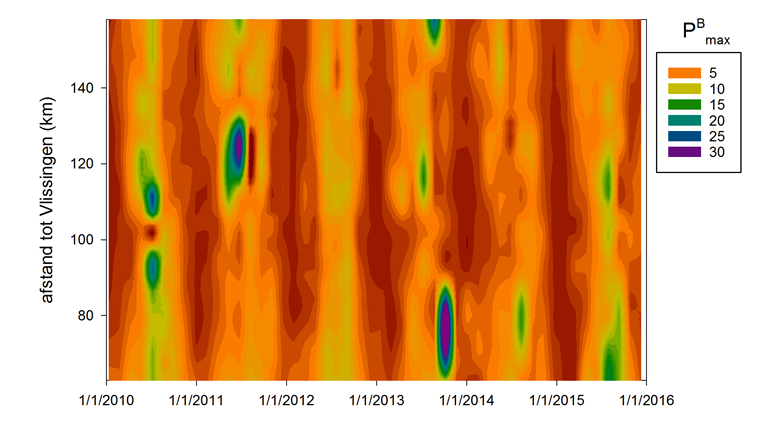
Figuur 6-15 contourplot PBmax. De waarden zijn gemaximaliseerd op 30 mgC (mg (chla)-1h-1).
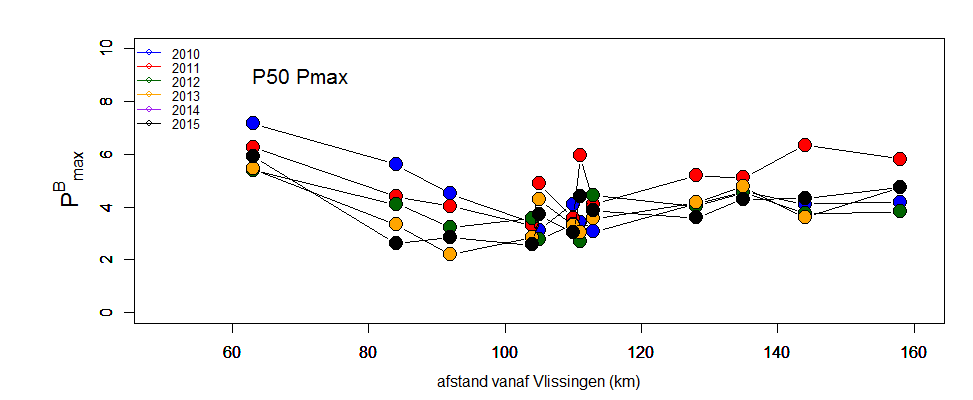
Figuur 6-16 Mediane waarden PBmax.
αB.
De initiële helling van de PI-curve (2.6.4), αB, geeft informatie over de affiniteit voor licht bij lage lichtintensiteiten. Een hoge waarde van αB betekent dat de algen een hoge fotosynthese efficiëntie hebben bij laag licht, en dat is karakteristiek voor algen die zich hebben aangepast aan een milieu met een gemiddeld lage beschikbaarheid van licht. In Figuur 6-17 is de variatie voor de verschillende stations en jaren weergegeven in een boxplot.
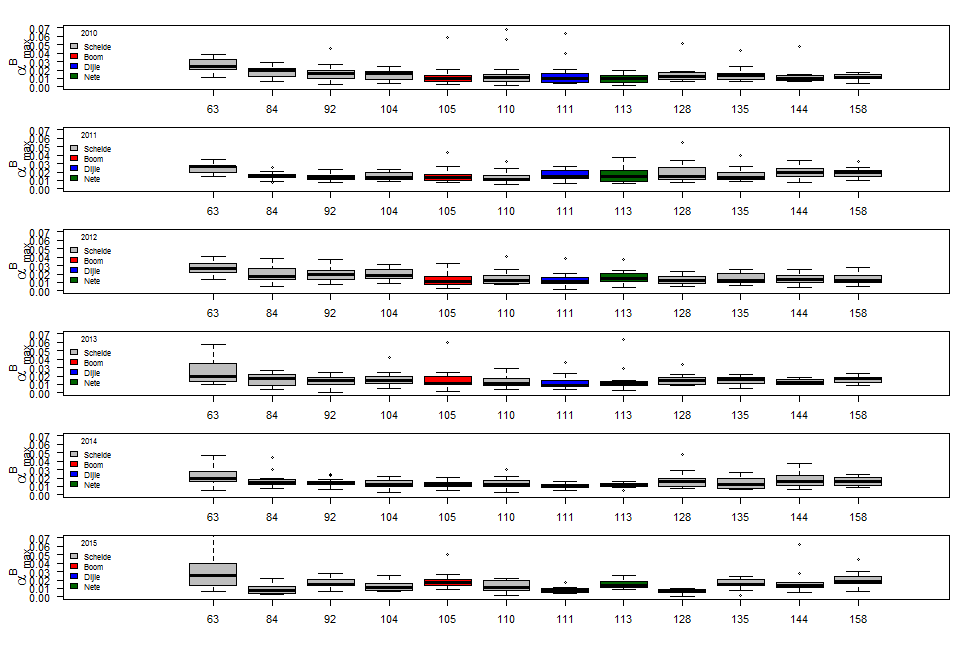
Figuur 6-17 overzicht van de variatie in αB. Op de X-as de afstand tot Vlissingen (km)
In het algemeen zijn de waarden het laagst rond Temse/Lippenbroek en het hoogst bij de grens. Richting Melle neemt αB in 2014 en 2015 weer enigszins toe, maar er zijn ook jaren dat er weinig ruimtelijke variatie is in αB.
In vorige rapportages is aangetoond dat PBmax en αB lineair en positief aan elkaar gekoppeld zijn, wat ook na het toevoegen van de data van 2015 het geval is (r2 = 0.62, p<0.001). Dit is ook zichtbaar als het verloop in αB gedurende het seizoen wordt geplot (zie Figuur 6-18 voor een 3-tal voorbeelden). Net als PBmax worden de maximale waarden in de zomer waargenomen. Op zich is dit opvallend omdat αB vooral bepaald wordt door de kans om licht in te vangen, en het fysische proces van lichtabsorptie is nauwelijks temperatuur afhankelijk. Verder valt op dat de maximale waarden in Zandvliet later in het jaar zijn waar te nemen dan in de overige stations
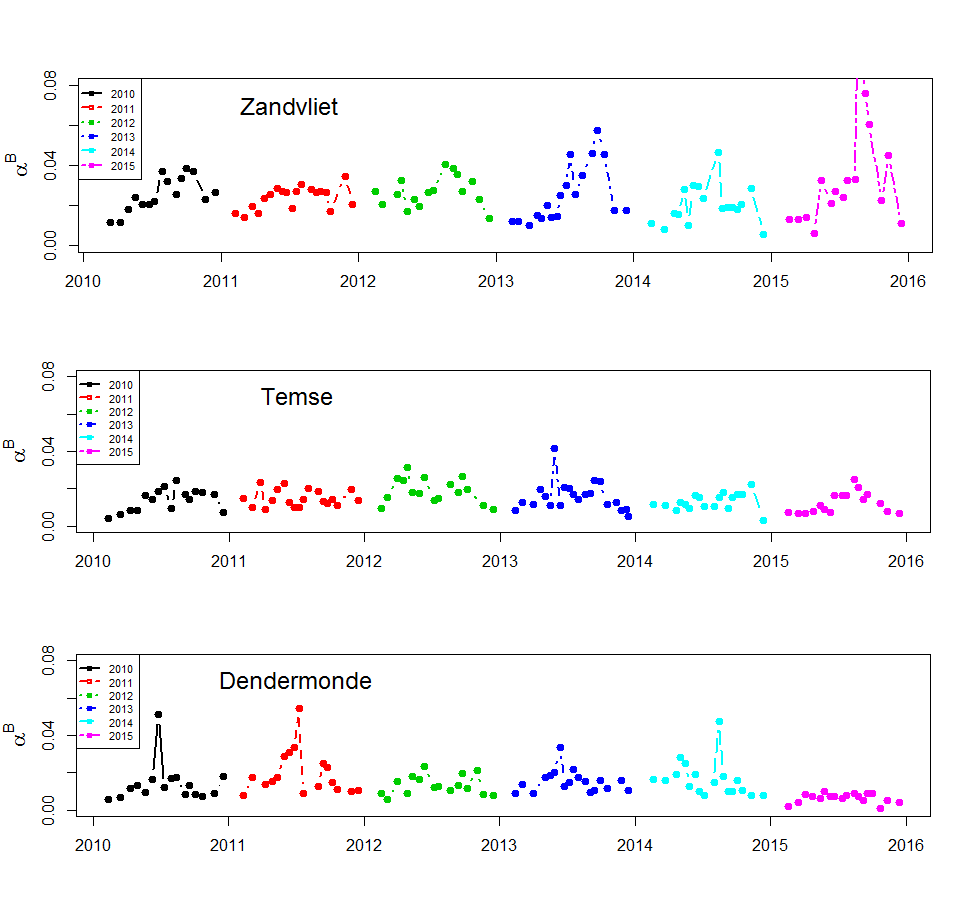
Figuur 6-18 Variatie in αB gedurende het seizoen voor 3 stations.
Ik
Ik (=PBmax/αB) is een maat voor de lichtintensiteit (I) waarbij de fotosynthese overgaat van lichtbeperkt naar beperking in de donkerreacties (Calvin cyclus). De overgang van licht-gelimiteerde fotosynthese activiteit naar lichtverzadigde fotosynthese (en waar de fotosynthesesnelheid dus gelimiteerd wordt door de snelheid waarmee CO2 enzymatisch gefixeerd kan worden) vindt namelijk plaats rond de waarde van Ik=PBmax/αB. In de jaren 2010 t/m 2013 neemt Ik toe van de grens tot aan Lippenbroek estuarien (km 110), en bovenstrooms van dit station lijken de waarden ongeveer constant te blijven (Figuur 6-19). Echter, in 2014 neemt Ik in Uitbergen en Melle weer af, wat op een mogelijk slechter lichtklimaat kan duiden. In 2015 is er vrij veel variatie in αB. In 2015 zijn er echter vrij veel baggerwerken geweest opwaarts Melle, wat mogelijk een invloed heeft gehad. Dit zal de komende jaren nader bekeken worden.
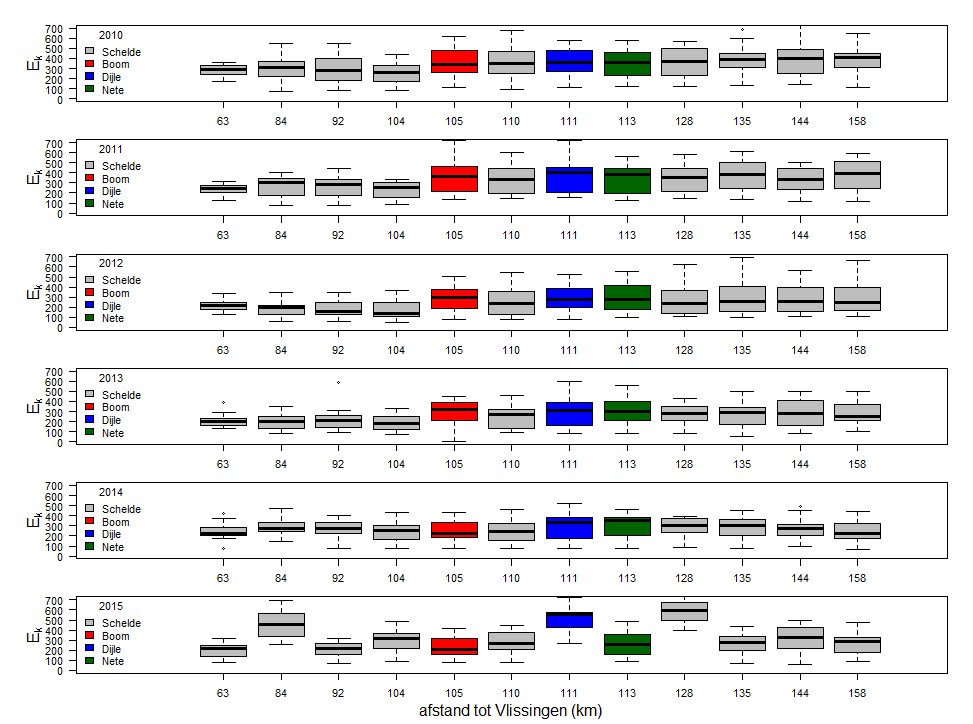
Figuur 6-19 boxplot Ik-waarden voor de verschillende stations
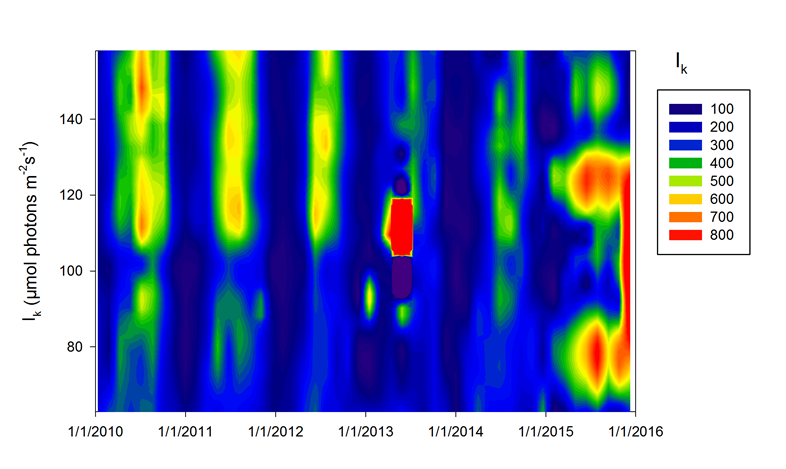
Figuur 6-20 contourplot Ik-waarden
De contourplot voor Ik (Figuur 6-20) lijkt te suggereren dat er een langzame afname in Ik plaatsvindt tussen 2010 en 2015, die tot stand komt in 2015. Dit jaar vertoont een vreemd patroon met hoge Ik-waarden rond km 75 en 120 in de zomer en aan het einde van het jaar.
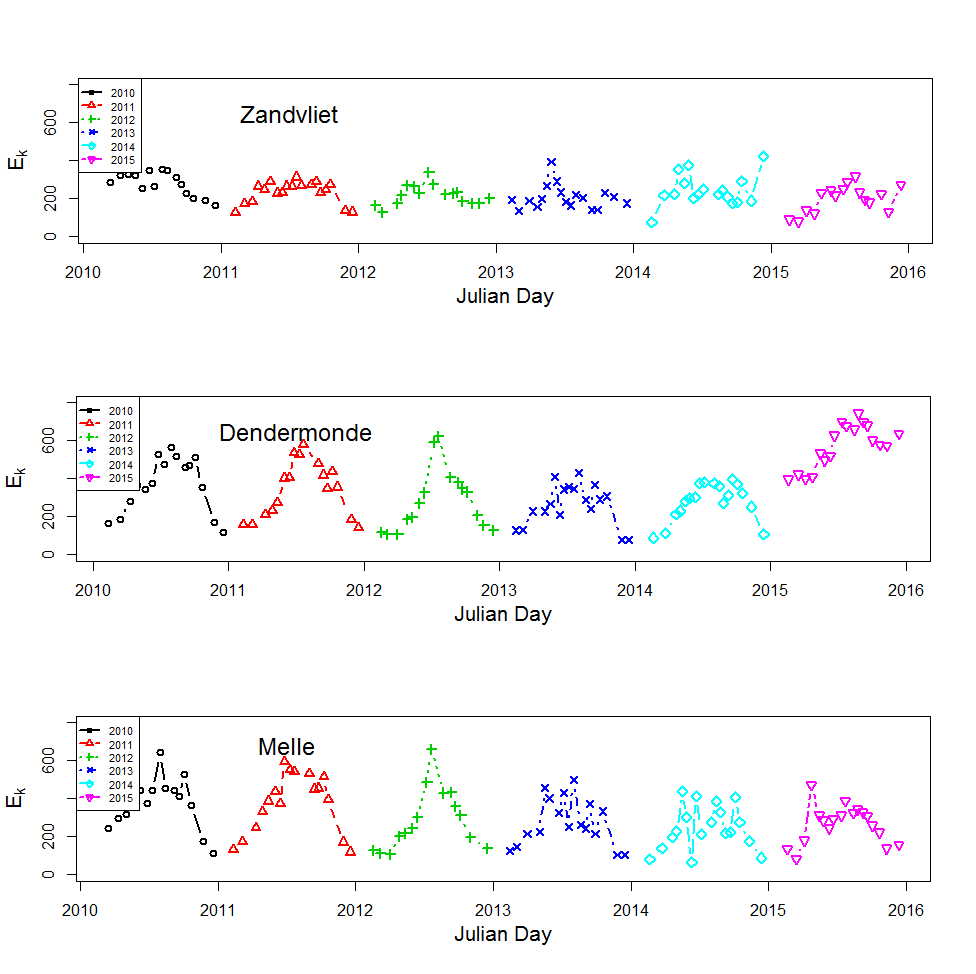
Figuur 6-21 verandering in Ik (µmol fotonen m-1 s-1) gedurende het seizoen
Deze afname in Ik per station is duidelijk zichtbaar als de Ik waarden worden geplot gedurende de verschillende jaren (Figuur 6-21). Deze afname lijkt in Melle gestopt in 2015. Dendermonde heeft duidelijk een afwijkend patroon in 2015. Of dit een artefact is in de C14-meting is onduidelijk maar kan niet geweten worden aan mogelijk foute chl a waarden.
6.3.3.Primaire productie
In Figuur 6-22 en Figuur 6-23 worden de dagproducties weergegeven in respectievelijk de Zeeschelde en de zijrivieren.
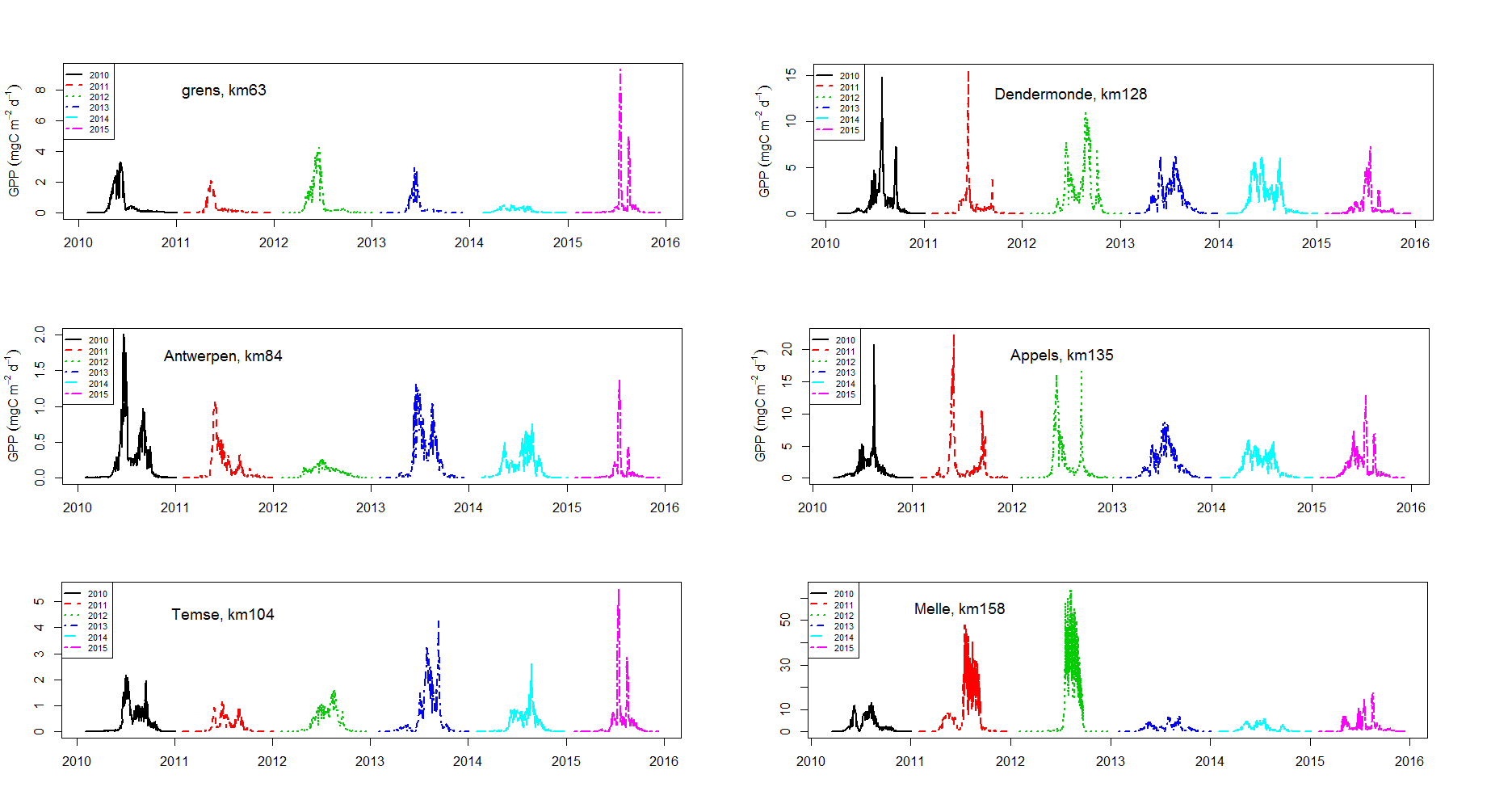
Figuur 6-22 Dagproducties voor een aantal stations in de Zeeschelde. Let op de verschillende Y-as schalen: het laagst voor Antwerpen en het hoogst voor Melle.
Wat opvalt, is de grote variatie van jaar tot jaar. Bij de grens zijn er in 2014 zeer lage dagproducties, terwijl dat voor de overige stations (met uitzondering van Melle) niet het geval was. Ook dicht bij elkaar gelegen stations kunnen flinke verschillen in patronen laten zien: zo zijn in Melle in zowel 2013 als 2014 de dagproducties gering, terwijl dit bij Appels (en Uitbergen), slechts 23 km (14 km) stroomafwaarts gelegen, niet duidelijk het geval was. Dit betekent dat lokale verschillen grote invloed kunnen hebben op de dagproducties. Een factor die hierbij een rol kan spelen is de getij gebonden variatie in de lichtextinctiecoëfficiënt kd. Eenzelfde grote variatie is waarneembaar in de zijrivieren (Figuur 6-23).
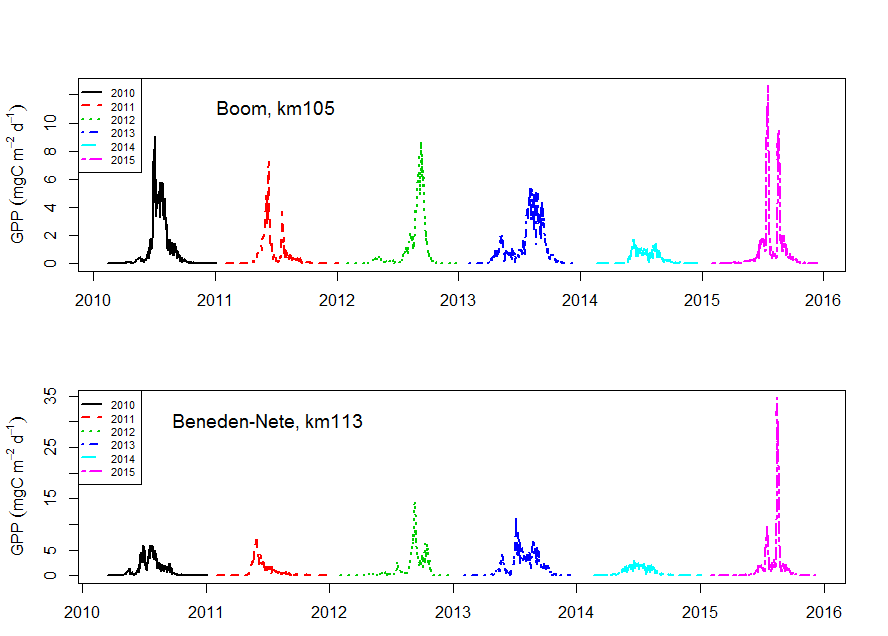
Figuur 6-23 Dagproducties in de zijrivieren de Rupel (bij station Boom) en in de Beneden-Nete.
De jaarproducties zijn weergegeven in Tabel 6-1 en Figuur 6-24. De jaarproducties nemen stroomopwaarts sterk toe. Vanaf Dendermonde (km128) is de toename stroomopwaarts gering, met uitzondering van Melle (km158) in 2011 en 2012, waarbij de jaarproducties aanzienlijk hoger waren dan in de overige jaren. De jaarlijkse variatie in de Beneden Zeeschelde lijkt beperkt maar dat is een gevolg van de lage producties. Als de standaardfout (standaarddeviatie als percentage van de gemiddelde jaarproducties in de periode 2010 t/m 2015) wordt uitgerekend dan zien we dat de variatie rondom het gemiddelde van 40 ± 15% bedraagt (Tabel 6-2).
Tabel 6-1 Jaarproducties (g C m-2jaar-1) voor de verschillende stations in de Zeeschelde en tijgebonden zijrivieren.
| Km | station | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 |
| 63 | Grens (Zandvliet) | 151 | 88 | 186 | 84 | 84 | 161 |
| 84 | Antwerpen | 81 | 49 | 27 | 78 | 51 | 25 |
| 92 | Kruibeke (Hoboken) | 88 | 66 | 41 | 75 | 74 | 69 |
| 104 | Temse | 124 | 67 | 113 | 181 | 206 | 234 |
| 105 | Boom | 305 | 181 | 302 | 361 | 122 | 313 |
| 110 | Lippenbroek | 492 | 147 | 491 | 963 | 279 | 661 |
| 111 | Dijle | 307 | 705 | 260 | 432 | 153 | 560 |
| 113 | Nete | 345 | 273 | 411 | 549 | 220 | 544 |
| 128 | Dendermonde | 721 | 437 | 1205 | 910 | 1014 | 485 |
| 135 | Appels | 794 | 1035 | 1182 | 1108 | 1117 | 999 |
| 144 | Uitbergen | 1027 | 699 | 1507 | 1267 | 669 | 1826 |
| 158 | Melle | 1679 | 4183 | 4757 | 987 | 960 | 1403 |
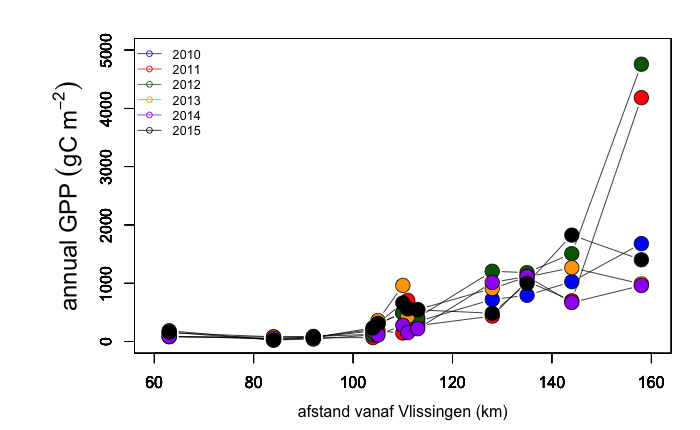
Figuur 6-24 Variatie in jaarproductie voor de verschillende stations in de periode 2010-2015.
Tabel 6-2 gemiddelde jaarproducties en standaarddeviatie (stdev) en standaardfout daarin.
| Km | station | gemiddeld | Stdev | st. error (%) |
| 63 | Grens (Zandvliet) | 125.7 | 45.7 | 36.3 |
| 84 | Antwerpen | 51.8 | 24.1 | 46.6 |
| 92 | Kruibeke (Hoboken) | 68.8 | 15.6 | 22.7 |
| 104 | Temse | 154.2 | 63.2 | 41.0 |
| 105 | Boom | 264.1 | 91.7 | 34.7 |
| 110 | Lippenbroek | 505.4 | 287.6 | 56.9 |
| 111 | Dijle | 402.8 | 204.4 | 50.7 |
| 113 | Nete | 390.4 | 137.2 | 35.1 |
| 128 | Dendermonde | 795.3 | 303.1 | 38.1 |
| 135 | Appels | 1039.1 | 136.4 | 13.1 |
| 144 | Uitbergen | 1165.9 | 457.6 | 39.2 |
| 158 | Melle | 2328.1 | 1690.5 | 72.6 |
6.3.4.PB and PB:B-ratios
Algen, en meer bijzonder kiezelwieren vormen de basis van de voedselketen en zijn dus essentieel voor het estuariene ecosysteem. Maar een teveel aan algen kan leiden tot een problematische zuurstofvraag bij afsterven. Daarom is een evaluatie van algenbloei opgenomen in de Evaluatiemethodiek. Welke concentratie aan algen nu ideaal is, en vanaf welke drempelwaarde er problemen kunnen rijzen, hangt af van tal van factoren. Het was daarom niet mogelijk een ondubbelzinnig criterium uit te werken voor de algenconcentratie. De Evaluatiemethodiek gaat ervan uit dat algen geen probleem vormen, zolang ze maar vlot opgegeten worden. Hiertoe werden de PB en PB:B ratio geselecteerd als rekenparameter.
De PB ratio (mgC (mg chla)-1 d-1) geeft informatie over de primaire productie per eenheid biomassa, hier benaderd aan de hand van de chl a concentratie. De ratio wordt berekend door de dagproductie (mgC m-2 d-1) te delen door het chl a gehalte over de gehele waterkolom (mg chl a m-2 = mg chl a m-3 x gemiddelde diepte (m)). De evaluatiemethodiek vereist daarnaast ook de berekening van de PB:B-ratio, die verkregen wordt door de PB-ratio te delen door de chl a concentratie (mg chl a m-3, waardoor de eenheid mgC (mg chla)-1 d-1 (mg chla m-3) ontstaat. Deze ratio geeft de verhouding weer tussen de primaire productie (per eenheid biomassa) en de biomassa aan algen. Een hoge ratio is ideaal, dit betekent immers dat de algen zeer productief zijn maar toch beperkt zijn in biomassa. Dit kan als de algen vlot opgegeten worden en er dus een goede doorstroming is van massa/energie in de voedselketen.
De variatie in de PB-ratio is weergegeven in een boxplot in Figuur 6-24 en in Tabel 6-3. De Y-as waarden zijn log-getransformeerd om beter de variatie in de mediane waarden te laten zien. De mediane waarde is ongeveer 2.8 mg C (mg chl a)-1 om dan af te nemen tot ~ 1 mg C (mg chl a)-1 voor de stations Antwerpen-Temse. Daarna neemt de mediane PB-ratio weer toe tot ongeveer 3 mg C (mg chl a)-1 bij Appels om vervolgens snel te stijgen tot meer dan 9 bij Melle.
De PB:B-ratio vertoont een ander beeld en bereikt minimale waarden bij Appels en Uitbergen en de hoogste waarde bij de grens. Dit wijst erop dat nabij de grens de algen productief zijn, maar dat er geen grote biomassa wordt opgebouwd, wellicht omdat de algen snel worden begraasd. Bij Appels en Uitbergen ligt de productie hoger, maar worden ook veel hogere biomassa’s opgebouwd. Dit is een indicatie dat de begrazing daar ondermaats is. De P90-waarden variëren van 8.4 bij de grens tot ~ 3 tussen Antwerpen en Kruibeke om vervolgens toe te nemen tot 9 respectievelijk 39 bij stations Uitbergen en Melle.
Als wordt aangenomen dat de C:Chl ratio ~ 40 mg:mg is, kan de bruto populatie groeisnelheid worden uitgerekend. Dit is bruto omdat geen rekening gehouden wordt met respiratoire verliezen in het donkere deel van de waterkolom en de nacht. De mediane groeisnelheden variëren van 0.03 dag-1 bij Antwerpen tot 0.24 bij Melle. De P90-waarden voor de groeisnelheden variëren tussen 0.1 en 0.3 met als uitzondering 0.9 voor Melle. Dit betekent dat de ongeveer maximale groeisnelheden (de P90-waarden) leiden tot cel verdubbelingstijden variëren van 9 tot 1 dagen, wat aantoont dat het fytoplankton zelfs als de omstandigheden het gunstigst zijn in het jaar toch nog meerdere dagen nodig hebben voor een celdeling, met uitzondering van Melle, waar ongeveer 1 verdubbeling per dag kan worden gerealiseerd. Let wel: deze waarden zijn een flinke onderschatting van de werkelijke verdubbelingstijd omdat zoals gezegd geen rekening is gehouden met de ademhalingsverliezen in het donker.
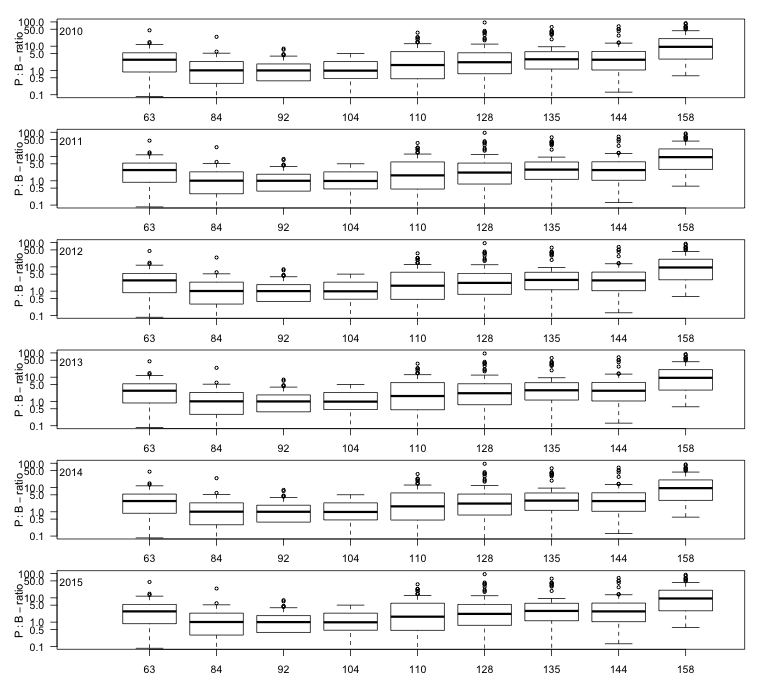
Figuur 6-25 Boxplot van de PB (P:B) waarden van het fytoplankton voor de verschillende stations. Let op: de Y-as is log-getransformeerd om de verschillen duidelijker zichtbaar te maken.
Tabel 6-3 PB- en PB:B-ratio’s en “bruto populatie groeisnelheden” onder aanname van een C:Chla ratio van 40.
| P50-waarden | P90-waarden | |||||
| km | PB | PBB | Growth rate | PB | PBB | Growth-rate |
| 63 | 2.77 | 0.47 | 0.07 | 8.37 | 2.60 | 0.21 |
| 84 | 1.08 | 0.15 | 0.03 | 3.90 | 0.70 | 0.10 |
| 92 | 1.01 | 0.11 | 0.03 | 2.95 | 0.42 | 0.07 |
| 104 | 1.01 | 0.10 | 0.03 | 3.30 | 0.30 | 0.08 |
| 110 | 1.68 | 0.09 | 0.04 | 12.24 | 0.83 | 0.31 |
| 128 | 2.21 | 0.06 | 0.06 | 11.48 | 0.53 | 0.29 |
| 135 | 2.92 | 0.05 | 0.07 | 9.44 | 0.71 | 0.24 |
| 144 | 2.92 | 0.05 | 0.07 | 9.32 | 0.56 | 0.23 |
| 158 | 9.44 | 0.23 | 0.24 | 38.72 | 1.13 | 0.97 |
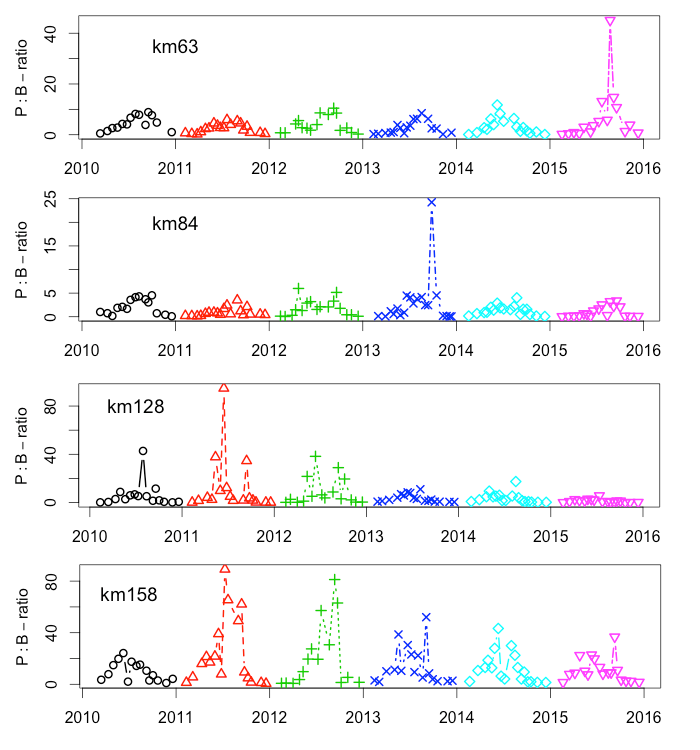
Figuur 6-26 Verloop in PB (P:B) -ratio voor een aantal stations (Grens (Zandvliet), Antwerpen, Dendermonde, Melle).
Het verloop in het groeiseizoen laat zien dat zoals verwacht mag worden de maximale PB-ratio’s in de zomer worden waargenomen, maar dat er grote variaties zijn tussen de verschillende jaren. Tussen Dendermonde en Melle lijkt de PB-ratio af te nemen van 2013, maar of er sprake is van een trend kan wegens gebrek aan data voor meerdere jaren nog niet worden gezegd omdat de waarden van 2010 ook vrij laag zijn in vergelijkbaar met die van 2014 en 2015. De extra hoge PB-ratio’s in 2015 voor km63 en in 2014 bij km84 zouden te wijten kunnen zijn aan relatief lage chl a concentraties.
In Figuur 6-27 en Figuur 6-28 zijn de contourplots van respectievelijk de PB-ratio en de PB:B-ratio opgenomen. De PB-ratio laat het verwachte verloop zien met de hoogste ratio’s in de zomer en stroomopwaarts.
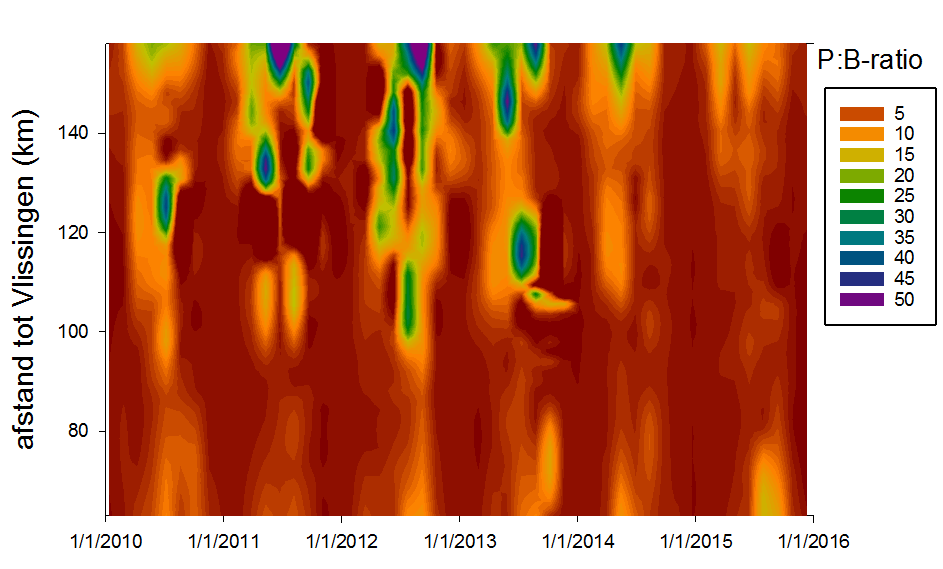
Figuur 6-27 contourplot PB-ratio (mg C mg-1 chla dag-1)
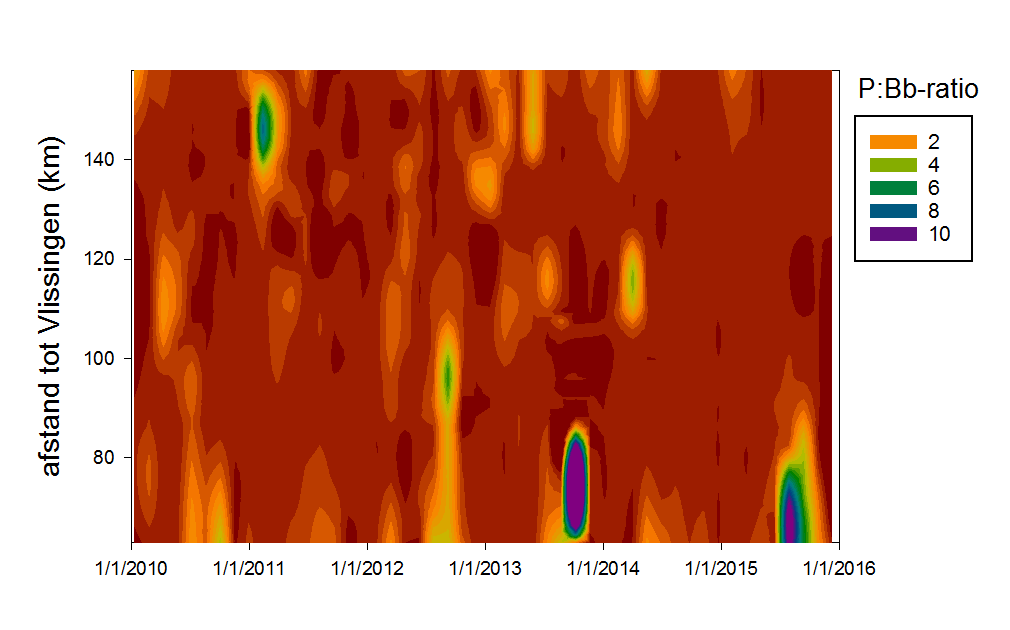
Figuur 6-28 contourplot PB:B-ratio (mg C mg-1 chla dag-1 (mg chla m-3))
De PB ende PB:B-ratio lijken niet met elkaar gecorreleerd, zoals hierboven al opgemerkt. De hoogste PB:B-ratio’s worden gevonden in het gebied tussen de grens en Antwerpen, daar waar het systeem zeer troebel is.
6.4.Fytoplankton anno 2015
6.4.1.Biomassa
De fytoplankton biomassa (benaderd via chl a) neemt stroomopwaarts toe van het brakke water bij Boei 87 tot het zoete water bij Melle (Figuur 6-29). Ook de standaarddeviatie neemt stroomopwaarts toe, dit komt doordat er een grotere variatie is in fytoplankton biomassa over het jaar. Dit is ook zichtbaar in Figuur 6-30 waar vanaf april stroomopwaarts de fytoplanktonbloei start en zich stroomafwaarts verspreidt. Chl a waardes van +/- 300 µg/l worden in mei/juni en in augustus gemeten bij de stations stroomafwaarts.
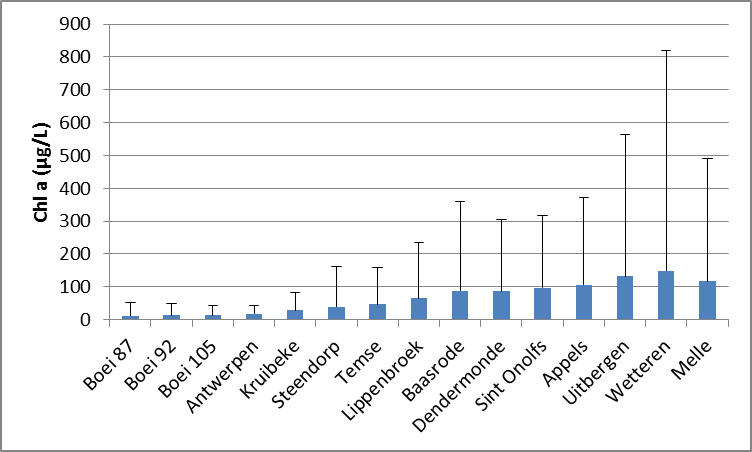
Figuur 6-29 Gemiddelde Chl a waarden van 2015 met standaarddeviatie voor de meetstations van het Schelde estuarium (stroomopwaarts van Boei 87 tot Melle).
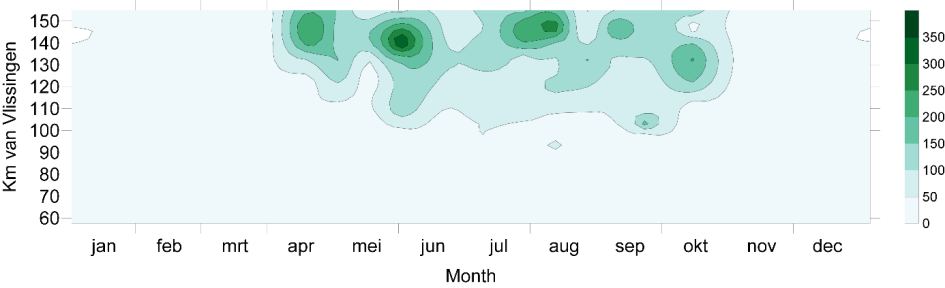
Figuur 6-30 Chl a concentraties (µg l-1) van het brakke deel bij de Nederlands-Belgische grens (± 60 km) tot aan het zoetwater deel bij Gent (± 160 km) van het Schelde estuarium.
Net als in 2011 was er in 2015 een lagere fytoplankton biomassa in vergelijking met de andere jaren. 2011 en 2015 waren jaren met bijzonder veel zwevende stof in het water. Lichtlimitatie heeft wellicht de algenbloei beperkt. Sinds 2008 lijkt er een geleidelijke verschuiving van de maximale bloei intensiteit naar het bovenstroomse deel van het Schelde estuarium. Opvallend is ook het ontbreken van een sterke bloei in het voorjaar nabij de Grens.
6.4.2.Resultaten fytoplankton gemeenschapssamenstelling
De pigmentanalyse toont aan dat er een dominantie is van fucoxanthin (Figuur 6-31), wat een belangrijk pigment voor diatomeeën is. Dit indiceert dat de fytoplankton gemeenschap in 2015, net als voorgaande jaren, voornamelijk uit diatomeeën bestaat. Chl b and lutein zijn indicatief voor Chlorophyta (groenalgen) en euglenophytes, deze pigmenten zijn in lagere hoeveelheden aanwezig in vergelijking tot fucoxanthin. Hetzelfde geldt voor de pigmenten zeaxanthin, peridinin en alloxanthin, die respectievelijk kenmerkend zijn voor cyanobacteriën, dinophytes en cryptophytes (Figuur 6-31). De uitkomsten van de pigmentanalyse worden ondersteund door microscopische tellingen (Figuur 6-32), waaruit blijkt dat de belangrijkste groep diatomeeën zijn en de andere groepen in veel lagere aantallen voorkomen.
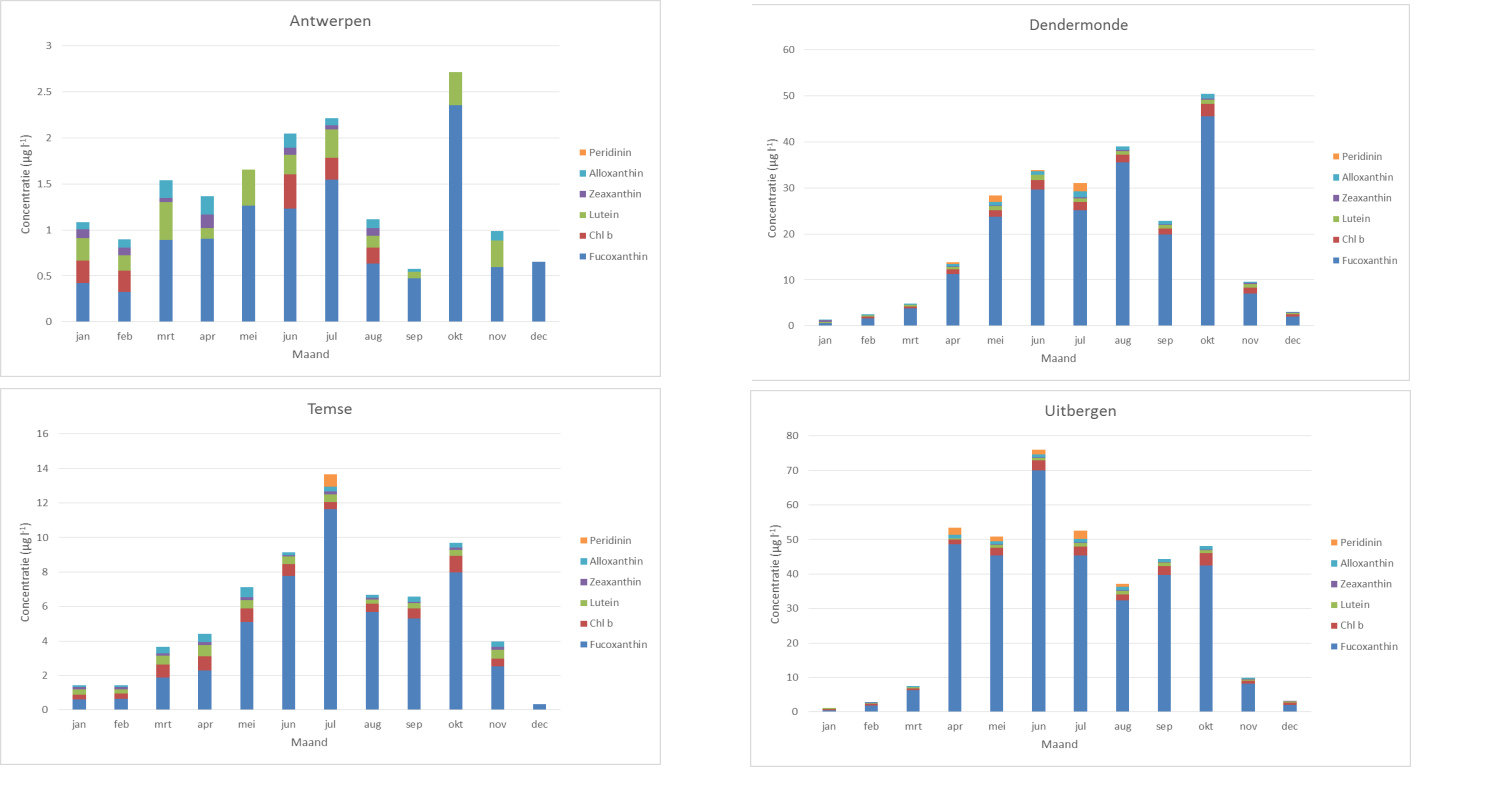
Figuur 6-31 Maandelijkse pigment samenstelling in 2015 voor de stations Antwerpen, Temse, Dendermonde en Uitbergen. Let op dat de verschillende figuren andere y assen hebben.
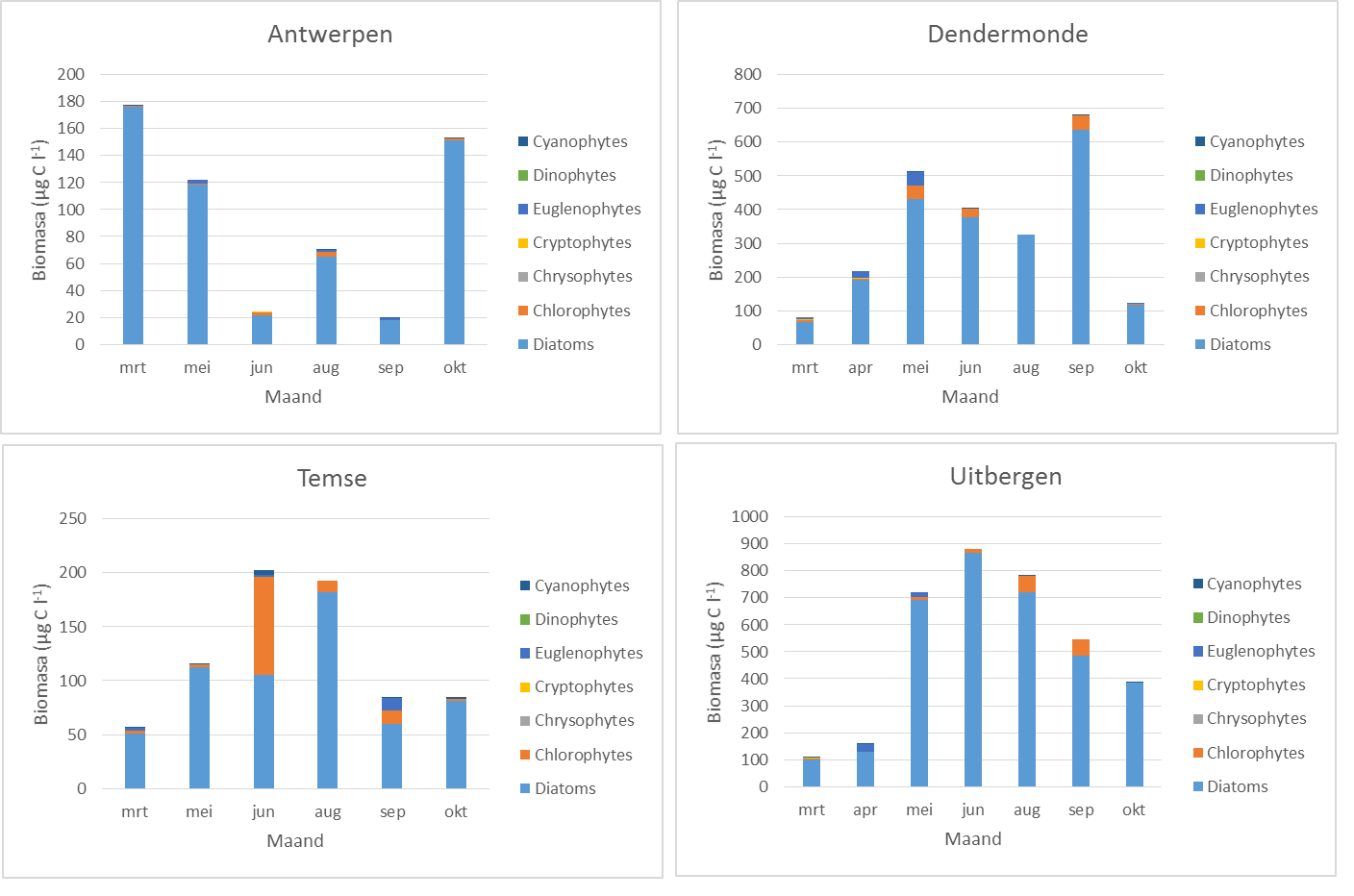
Figuur 6-32 Maandelijkse fytoplankton samenstelling (biomassa in µg C l-1) in 2015 voor de stations Antwerpen, Temse, Dendermonde en Uitbergen. Let op dat de verschillende figuren een andere y-as hebben.
De microscopische bepaling van de fytoplankton gemeenschap toont aan dat de diatomeen de belangrijkste groep zijn. Echter in juni is er een hoge biomassa van groenalgen, namelijk Pediastrum, Actinastrum, Crucigeniella en Scenedesmus. De diatomeeën gemeenschap bestaat voornamelijk uit Actinocyclus normannii, Thalassiosira en Cyclotella.
6.4.3.PB en PB:B-ratio
Beide ratio’s werden reeds besproken in 6.3.4 voor de voorbije 6 jaar. Beide parameters werden in de evaluatiemethodiek opgenomen om de algenbloei te evalueren in functie van energiedoorstroming. Volgende criteria worden gehanteerd:
PB en PB:B worden per zone bepaald per maand tijdens de periode maart tot en met september. Jaargemiddeld dienen per zone volgende minima gehaald te worden:
- Een PB – waarde van 10 en een PB:B van 1,5 in de zoete zone met korte verblijftijd.
- Een PB – waarde van 5 en een PB:B van 0,75 in de zone met sterke saliniteitsgradiënt, de oligohaliene zone en de zoete zone met lange verblijftijd.
Voor 2015 wordt enkel in de zone met sterke saliniteitsgradiënt voldaan aan deze criteria (Tabel 6-4), in de andere zones is de primaire productie te laag in verhouding tot het aantal algen. Dit kan wijzen op een slechte doorstroming in de voedselketen: de algen zijn weinig productief maar kunnen toch biomassa opbouwen. De evaluatie voor deze parameter is bijgevolg negatief. Het gemiddelde verloop van beide parameters is weergegeven in Figuur 6-33 en Figuur 6-34.
Tabel 6-4 gemiddelde waarde per zone voor PB en PB:B ratio voor de periode maart – september 2015 (2015) en het toetscriterium hiervoor uit de Evaluatiemethodiek (E.M.)
| zone | PB | PBB | ||
| E.M. | 2015 | E.M. | 2015 | |
|
sterke saliniteitsgradient |
5 | 5,6 | 0,75 | 2,50 |
|
oligohalien |
5 | 1,9 | 0,75 | 0,27 |
|
zoet lange verblijftijd |
5 | 2,4 | 0,75 | 0,11 |
|
zoet korte verblijftijd |
10 | 7,4 | 1,50 | 0,15 |
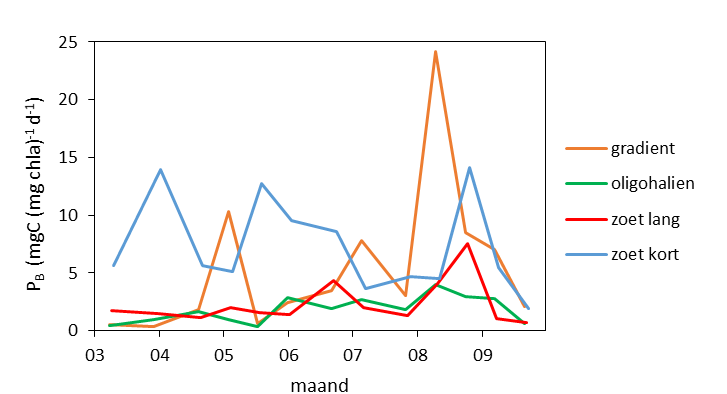
Figuur 6-33 gemiddelde PB ratio per maand per zone in de Zeeschelde voor 2015
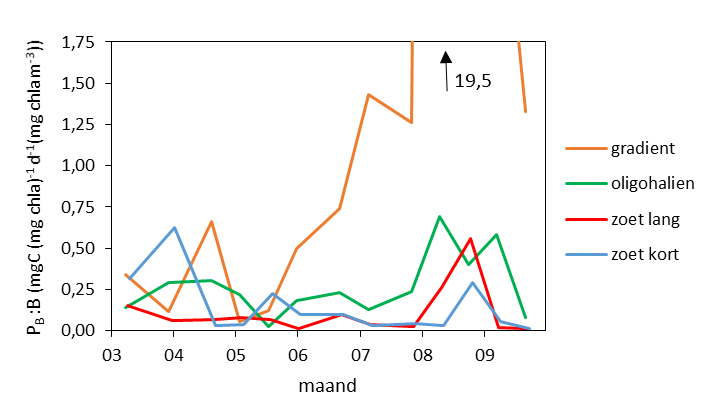
Figuur 6-34 gemiddelde PB:B ratio per maand per zone in de Zeeschelde voor 2015
6.4.4.Cyanobacteriën
Cyanobacteriën, blauwalgen of blauwwieren zijn strikt genomen geen algen, maar een groep binnen het domein van de bacteriën. Ze zijn wel, net als algen, in staat om aan fotosynthese te doen. Blauwalgen zijn berucht omdat ze bij grote bloei voor overlast kunnen zorgen. Sommige soorten blauwalgen kunnen immers voor de mens toxische stoffen uitscheiden. Ook voor dieren (vee) kunnen ze toxisch zijn. Daarom zijn in de Evaluatiemethodiek criteria opgenomen voor deze soort: vanaf een cellendichtheid van meer dan 2 x 107 cellen per liter is er sprake van problematische cyanobacteriebloei, maandelijks te bepalen in lente en zomer, in brak en zoet, per zone.
Tabel 6-5 geeft een overzicht van de cellendichtheid tijdens het groeiseizoen voor een selectie bemonsterde stations. Er is nergens sprake van een problematische bloei (max 5*106 cellen l-1). Hoe lager de saliniteit, hoe groter het aandeel cyanobacteriën, waarbij de dominante blauwalgen Planktotrix (Figuur 6-35) en Limnothrix (Figuur 6-36) zijn. Blauwalgen vormen dus nergens een probleem in de Zeeschelde: de evaluatie voor deze rekenparameter is positief.
De in het estuarium aanwezige cyanobacteriën worden waarschijnlijk vooral via de zijrivieren aangevoerd. Na droge perioden kan er na een stortbui een grote influx van cyanobacteriën optreden vanuit de toevoerrivieren wat eventueel tijdelijk een lokaal probleem zou kunnen vormen, maar tot op heden hebben we geen aanwijzingen voor een significante lokale ontwikkeling van blauwalgen in de Zeeschelde. Ook hier zijn lokale residentietijden van water wellicht cruciaal (naast licht, nutriënten etc uiteraard).
Tabel 6-5 Totaal aantal cyanobacteriën in het Schelde estuarium van maart t/m oktober 2015 (cellen l-1)
|
Station |
Km | mrt | apr | mei | jun | jul | aug | sep | okt |
|
Antwerpen |
78.5 | 2.5E+04 | 3.3E+04 | 0.0E+00 | 0.0E+00 | 9.8E+04 | 2.5E+04 | 3.5E+04 | 1.4E+05 |
|
Temse |
97.5 | 4.3E+05 | 4.5E+05 | 0.0E+00 | 0.0E+00 | 5.1E+05 | 0.0E+00 | 1.0E+06 | 3.3E+04 |
|
Dendermonde |
121.5 | 2.5E+05 | 5.1E+05 | 5.9E+05 | 6.4E+05 | 2.0E+05 | 0.0E+00 | 2.8E+05 | 4.5E+05 |
|
Uitbergen |
140 | 5.5E+05 | 9.3E+05 | 1.2E+06 | 0.0E+00 | 1.8E+06 | 3.0E+05 | 6.7E+05 | 3.8E+04 |
|
Bovenschelde |
180 | 1.7E+05 | 8.2E+04 | 5.3E+05 | 0.0E+00 | 4.6E+06 | 1.9E+06 | 4.6E+05 | 3.7E+06 |
|
Durme |
185 | 0.0E+00 | 0.0E+00 | 1.0E+06 | 1.9E+05 | 0.0E+00 | 0.0E+00 | 5.1E+05 | 0.0E+00 |
|
Dender |
190 | 2.1E+05 | 1.9E+06 | 2.0E+05 | 8.4E+05 | 0.0E+00 | 0.0E+00 | 6.5E+05 | 2.4E+05 |
|
Beneden-Nete |
Nete | 5.3E+05 | 1.1E+05 | 0.0E+00 | 8.6E+05 | 6.5E+05 | 0.0E+00 | 0.0E+00 | 0.0E+00 |
|
Boom (Rupel) |
Rupel | 5.9E+04 | 1.6E+05 | 1.9E+05 | 1.2E+06 | 0.0E+00 | 0.0E+00 | 7.5E+04 | 3.0E+05 |
|
Dijle |
Dijle | 2.0E+05 | 2.1E+05 | 0.0E+00 | 0.0E+00 | 9.3E+05 | 1.2E+06 | 0.0E+00 | 5.6E+05 |
|
Zandvliet |
- | 2.8E+04 | 0.0E+00 | 1.6E+04 | 2.8E+04 | 1.5E+05 | 1.1E+05 | 1.3E+05 | 9.2E+03 |
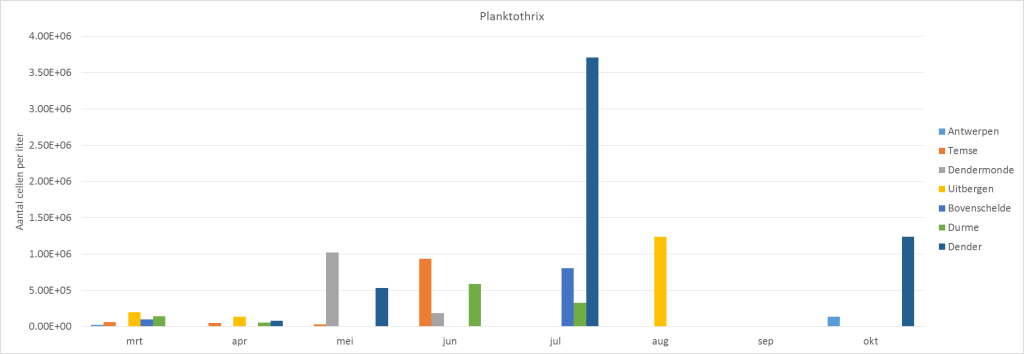
Figuur 6-35 Planktothrix in de Schelde van maart t/m oktober 2015 (cellen l-1). Antwerpen: 78.5, Temse: 97.5, Dendermonde: 121.5, Uitbergen: 140, Bovenschelde: 180, Durme 185 en Dender: 190 km van de uitmonding bij Vlissingen.
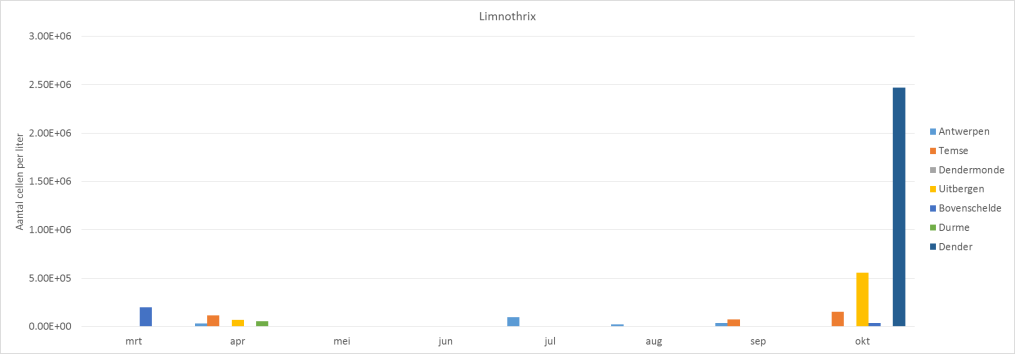
Figuur 6-36 Limnothrix in de Schelde van maart t/m oktober 2015 (cellen l-1). Antwerpen: 78.5, Temse: 97.5, Dendermonde: 121.5, Uitbergen: 140, Bovenschelde: 180, Durme 185 en Dender: 190 km van de uitmonding bij Vlissingen.
6.4.5.Phaeocystis
Wanneer de groenalg Phaeocystis in grote celdichtheden voorkomt, wordt ze als plaagalg beschouwd. Vanaf een cellendichtheid van meer dan 4 x 106 cellen per liter is er sprake van problematische Phaeocystis bloei. De algen vormen dan immers kolonies, die te groot zijn om nog door gegeten te worden door het meeste zoöplankton en benthos.
Tellingen wijzen uit dat in de Zeeschelde Phaeocystis niet in significante aantallen aanwezig is. Identificatie is wellicht wel bemoeilijkt door fragmentatie van kolonies maar op basis van de dichtheden van kleine flagellaten/eencelligen in de tellingen kan men besluiten dat Phaeocystis geen probleem vormt in de Zeeschelde. De evaluatie voor deze rekenparameters is bijgevolg positief.
6.5.Fytobenthos
6.5.1.Biomassa.
Binnen OMES wordt ook fythobenthos (= op de bodem levende algen) opgevolgd. Voor een overzicht van de staalname en staalnamelocaties, zie hoofdstuk 2.
De hoogste biomassa van fytobenthos, benaderd via chl a, wordt tijdens de maanden mei, juni, juli en augustus waargenomen met de hoogste waarden het meest stroomopwaarts te Appels (Figuur 6-37, Figuur 6-38). Er lijkt voor de zomerperiode een trend te zijn dat hoe verder stroomopwaarts van de Schelde hoe hoger de fytobenthos biomassa.
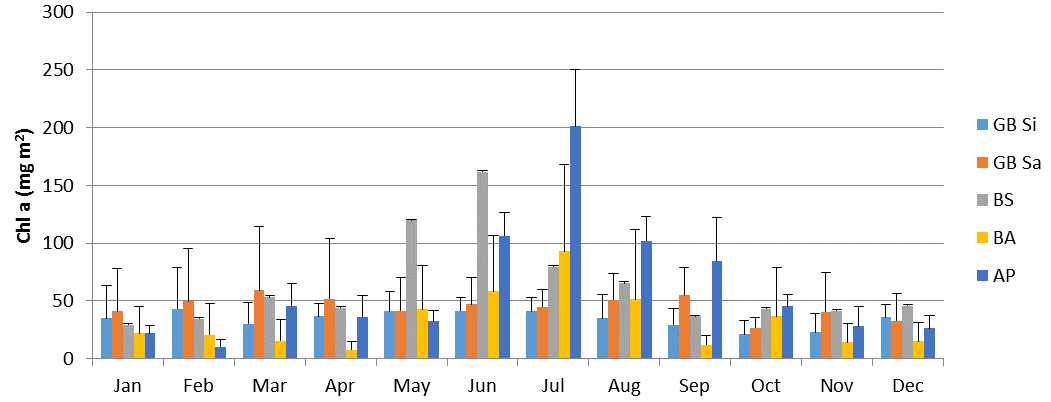
Figuur 6-37 Maandgemiddelden van de fytobenthos biomassa (Chl a in mg m2) van getijdeplaten langs het Schelde estuarium (GB Si: Groot Buitenschoor silt, GB Sa: Groot Buitenschoor sand, BS: Boerenschans, BA: Ballooi, AP: Appels).
6.5.2.Soortensamenstelling
Uit de pigmentanalyse blijkt dat de concentratie fuoxanthin een stuk hoger is dan zeaxanthin, lutein en Chl b (Figuur 6-39). Fucoxanthin wordt als indicator voor diatomeeën gebruikt, verwacht wordt dat dit de dominante groep is. Microscopische analyses van voorgaande jaren bevestigen deze resultaten. De Pennate diatomeeën van de genera Navicula, Cilindrotheca, Gyrosigma en Surirella waren dominant aanwezig op Groot Buitenschoor en Boerenschans. In de getijdengebieden van Ballooi en Appels werden de diatomeeen Caloneis amphisbaena, Actinocyclus normannii, Cylindrotheca sp. en de euglenophyte Euglena granulata voornamelijk gevonden. Een uitzondering van de dominantie van diatomeeën is tijdens de winteperiode voor de stations Ballooi en Appels.
Net als voorgaande jaren zijn Chl b en lutein in hogere concentraties aanwezig bij de stations stroomopwaarts, deze pigmenten zijn indicatief voor groenalgen en euglenophytes. Ze komen vooral voor rond de hoogwaterlijn waar de lichtcondities beter zijn. De indicatorpigment voor cyanobacteriën, zeaxanthin werd in heel 2015 niet in hoge concentraties waargenomen.
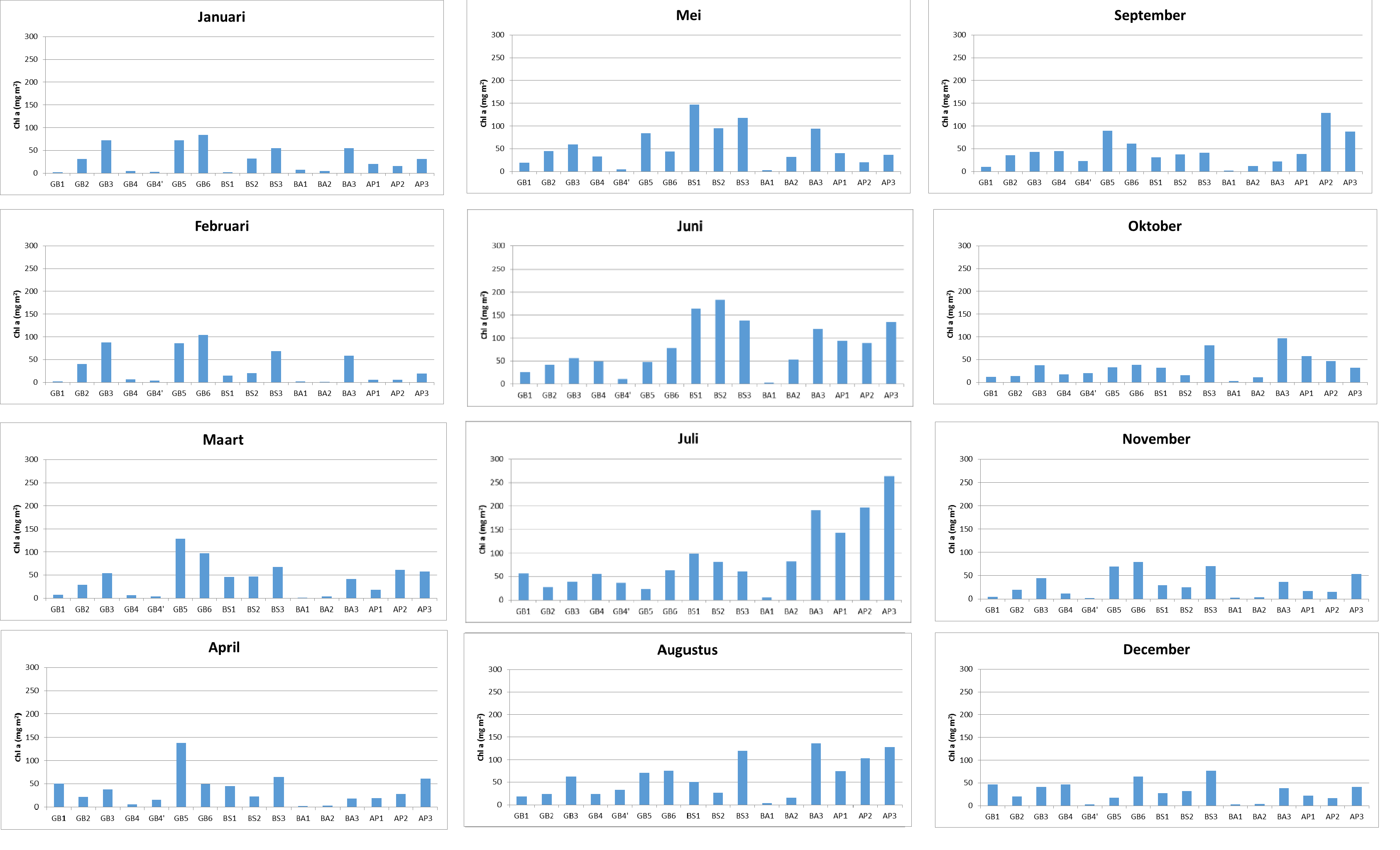
Figuur 6-38 ruimtelijk en tijdelijke verandering van fytobenthos biomassa in getijden gebieden van het Schelde estuarium (GB: Groot Buitenschoor, BS: Boerenschans, BA: Ballooi, AP: Appels).
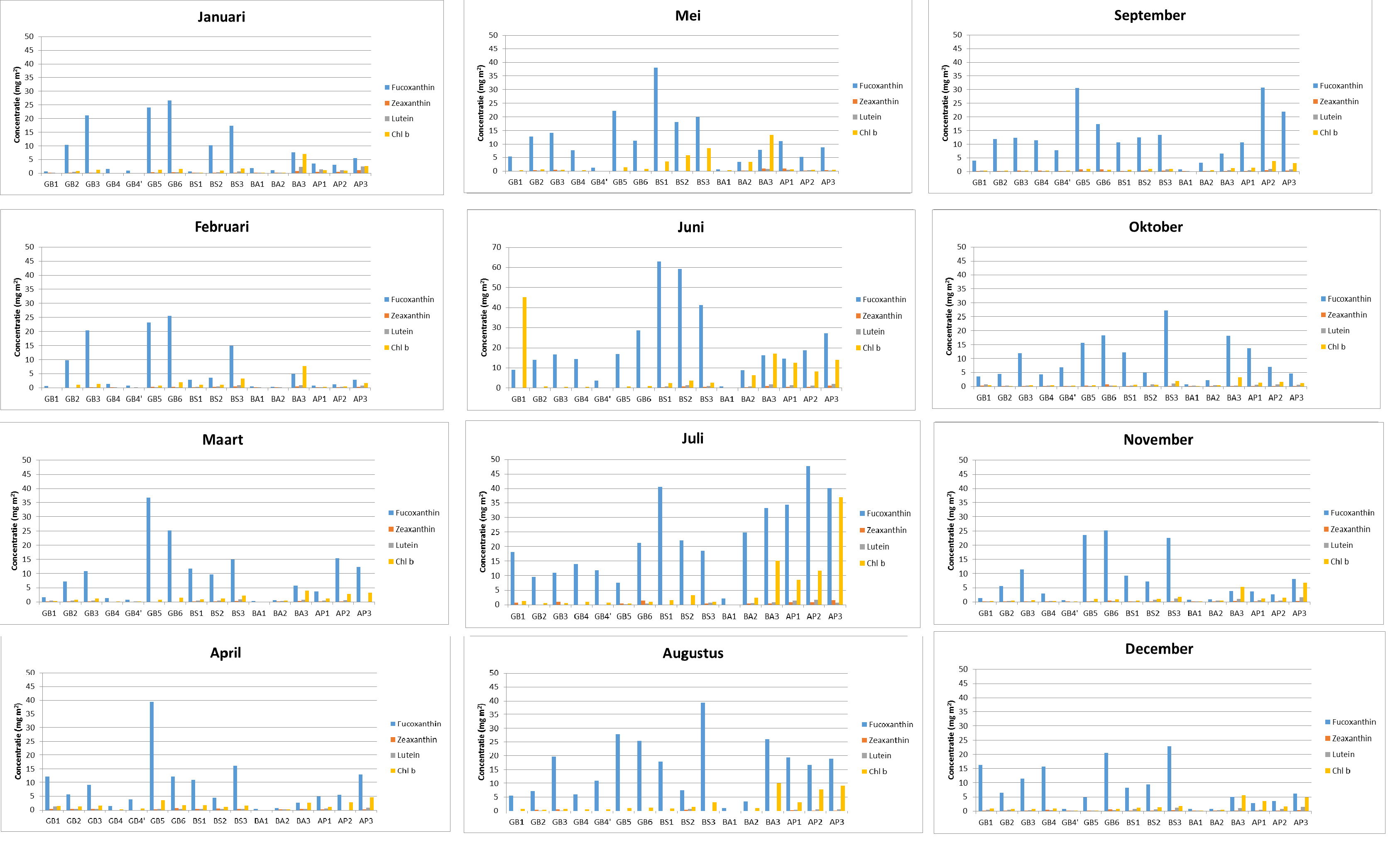
Figuur 6-39 Fytobenthos samenstelling in getijdengebieden van het Schelde estuarium (GB: Groot Buitenschoor, BS: Boerenschans, BA: Ballooi, AP: Appels). Let op: andere Y-as in juni.
6.5.3.Benthische primaire productie
De PBmax-waarden voor het microfytobenthos (MFB) variëren sterk per station en per jaar (Figuur 6-40). Vooral de stations op het slik van Ballooi laten een grote variatie van jaar tot jaar zien waarbij de hoogste PBmax-waarden werden waargenomen in 2012. Opvallend is verder dat de spreiding in de data voor de stations op Appels en in mindere mate in Boerenschans en Ballooi kleiner is dan die voor de stations op de 2 transecten op Groot Buitenschor.
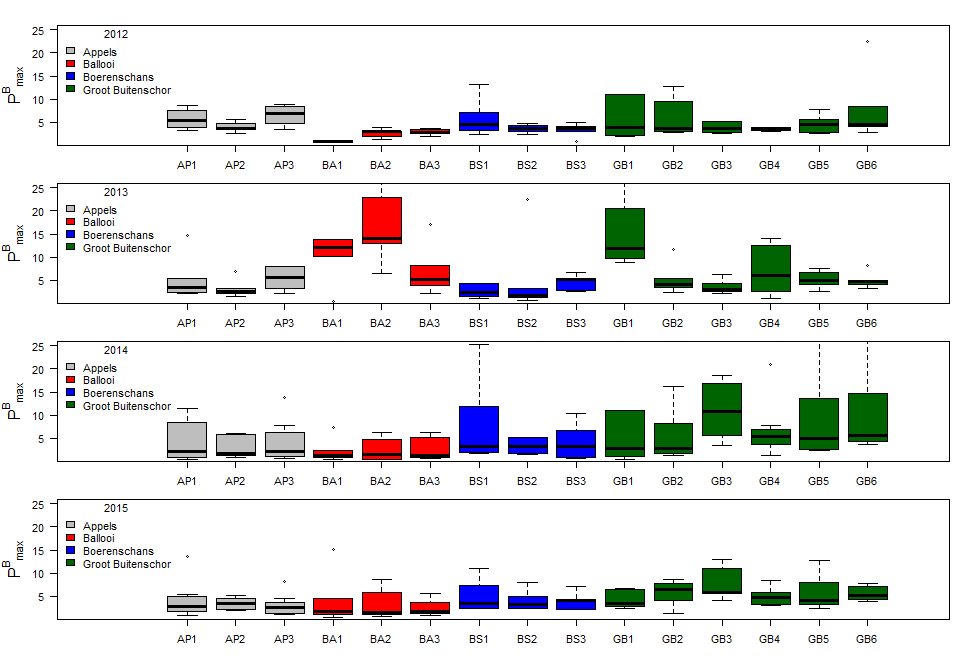
Figuur 6-40 boxplot van PBmax voor het microfytobenthos.
Als de PBmax-waarden worden geplot als functie van het seizoen lijkt en sprake van een voorjaarsbloei in 2013 en 2014 op de slikken van Appels en Ballooi (Figuur 6-41), terwijl in 2015 er sprake was van een zomerbloei. 2012 vertoont geen duidelijk patroon.
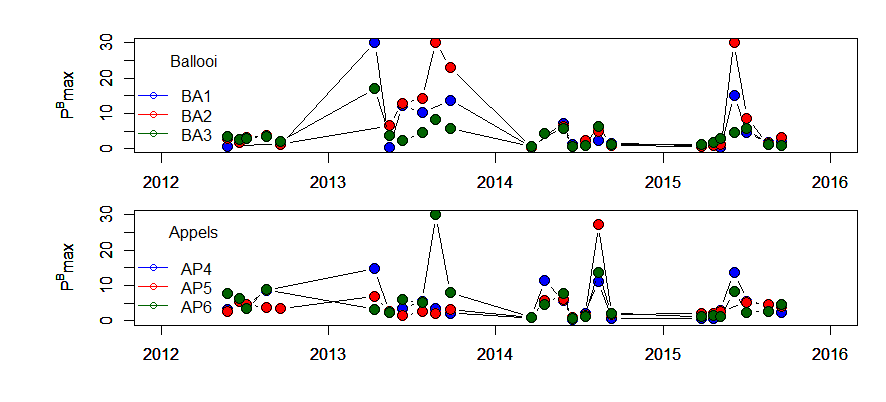
Figuur 6-41 timeplot PBmax-waarden voor de stations Ballooi en Appels
Deze voorjaars- en zomerbloei werd ook waargenomen in stalen van de slikken van Boerenschans en Groot buitenschor (Figuur 6-42). Voor de overige jaren was het patroon wat grilliger voor deze slikken.
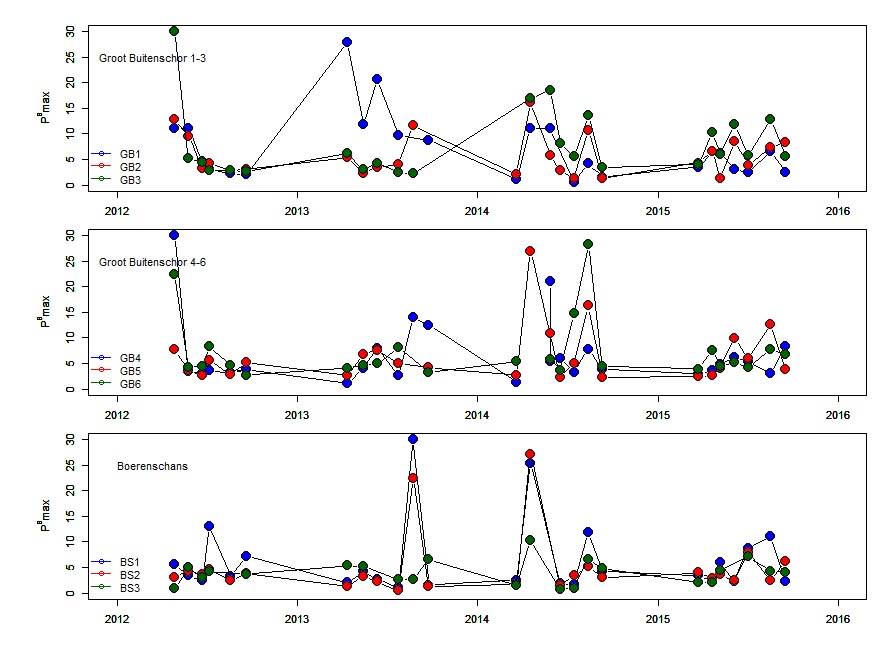
Figuur 6-42 timeplot PBmax-waarden voor de stations op de slikken van Groot Buitenschor (2 transecten) en Boerenschans.
Een factorial ANOVA liet op ln-getransformeerde PBmax-data toonde aan dat er geen significante verschillen tussen de jaren waren (p=0.137), maar dat er wel significante verschillen tussen de stations waren (p=0.00347). Er was geen significante interactie tussen jaar en station (p=0.593)
De variatie in de αB-waarden zijn geplot in boxplot (Figuur 6-43). De patronen in αB zijn vergelijkbaar met die van PBmax alhoewel de variatie van jaar tot jaar in vooral station Ballooi minder groot is. Opvallend is de variatie “skewed” verdeling van de αB-waarden in Appels in 2014. In het algemeen is de variatie in de αB-waarden het grootst van het MFB van de slikken van Groot Buitenschor. Ook hier liet een ANOVA zien dat er een significant verschil was tussen de stations (p<0.0001), maar dat er jaar (p=0.71) of interactie tussen jaar en station (p=0.355) niet significant waren.
De Ek waarden zijn te zien als boxplot in Figuur 6-44. Opvallend is dat de spreiding in de mediane waarden vrij beperkt is, vooral in 2012 en 2013, alhoewel in 2013 het MFB op stations GB3 en GB4 duidelijk lagere warden lieten zien, en datzelfde patroon is zichtbaar in 2015. Er is geen significant verschil in Ek tussen de verschillende station (p=0.253) of tussen de verschillende jaren (p=0.34).
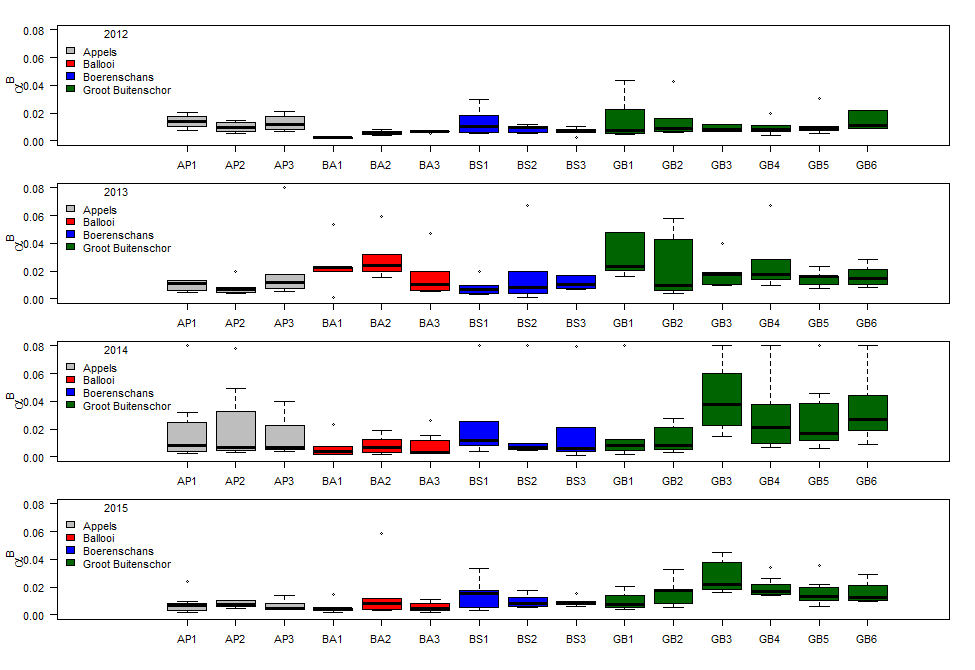
Figuur 6-43 boxplot αB waarden voor de verschillende stations en de verschillende jaren
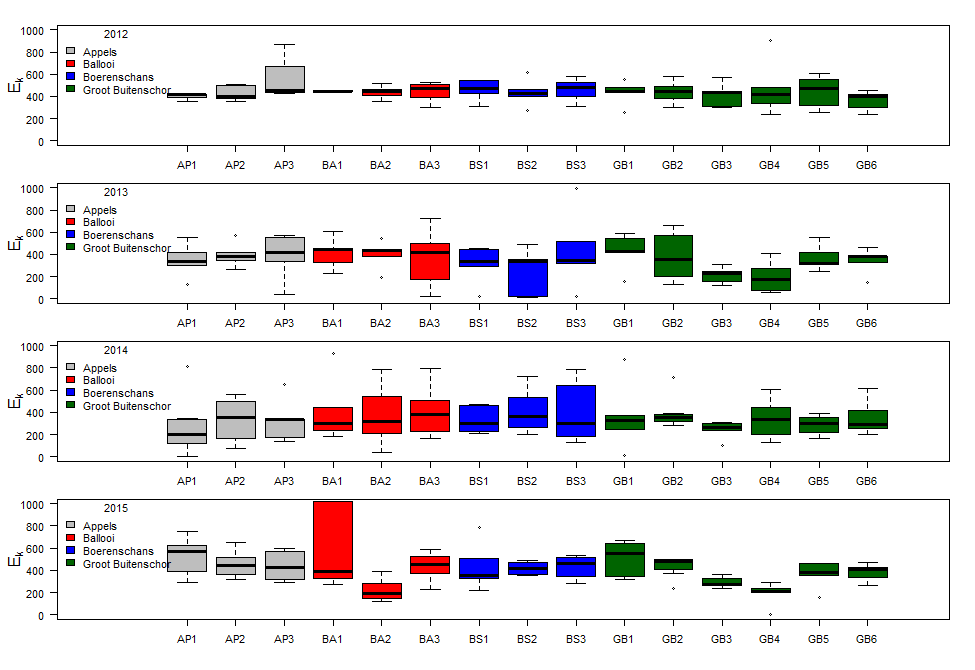
Figuur 6-44 boxplot Ek voor de verschillende stations en de jaren 2012 t/m 2015.
Als de mediane waarden voor de fitparameters tussen het fytoplankton en het microfytobenthos (MFB) worden vergeleken, dan valt op dat PBmax van het MFB gemiddeld 13% hoger is dan die van het fytoplankton, maar dat αB van het MFB gemiddeld 81% is van dat van het fytoplankton. Als gevolg is Ek van het MFB gemiddeld 37% hoger dan dat van het fytoplankton (Tabel 6-6). Dit lijkt er dus op alsof het MFB zich aanpast aan gemiddeld hogere lichtintensiteiten dan het fytoplankton, maar de verschillen zijn beperkt. De mediane PBmax van het MFB is 1.07-1.28 x zo groot als die van het fytoplankton.
Tabel 6-6 gemiddelde mediane waarden voor de fotosynthese fitparameters voor het microfytobenthos (MFB) en het fytoplankton
| Microfytobenthos | Fytoplankton | |||||
| Jaar | PBmax | αB | Ik | PBmax | αB | Ik |
| 2012 | 4.07 ± 1.11 | 0.0090 ± 0.0151 | 445 ± 26 | 3.82 ± 0.77 | 0.0151 ± 0.0045 | 235 ± 48 |
| 2013 | 4.87 ± 3.32 | 0.0128 ± 0.0064 | 351 ± 70 | 3.78 ± 0.93 | 0.0141 ± 0.0031 | 259 ± 49 |
| 2014 | 3.84 ± 2.46 | 0.0115 ± 0.0094 | 317 ± 46 | 3.77 ± 0.49 | 0.0138 ± 0.0026 | 273 ± 42 |
| 2015 | 4.34 ± 2.08 | 0.0125 ± 0.0033 | 390 ± 62 | 3.82 ± 0.98 | 0.0134 ± 0.0005 | 332 ± 132 |
| all | 4.28 ± 0.44 | 0.0114 ± 0.0017 | 378 ± 55 | 3.80 ±0 .03 | 0.0141 ± 0.0007 | 275 ± 41 |
6.6.Zooplankton
6.6.1.Abundantie en samenstelling zoöplankton gemeenschap
De resultaten worden voorgesteld per saliniteitszone: enerzijds het ‘brakwater traject’ de zone met sterke saliniteitsgradient station Grens (zone 4) en Antwerpen (zone5); en anderzijds de oligohaliene en zoete zone met stations Temse (zone 5) , Dendermonde (zone 6), Uitbergen (zone 7) en Melle (zone 7).
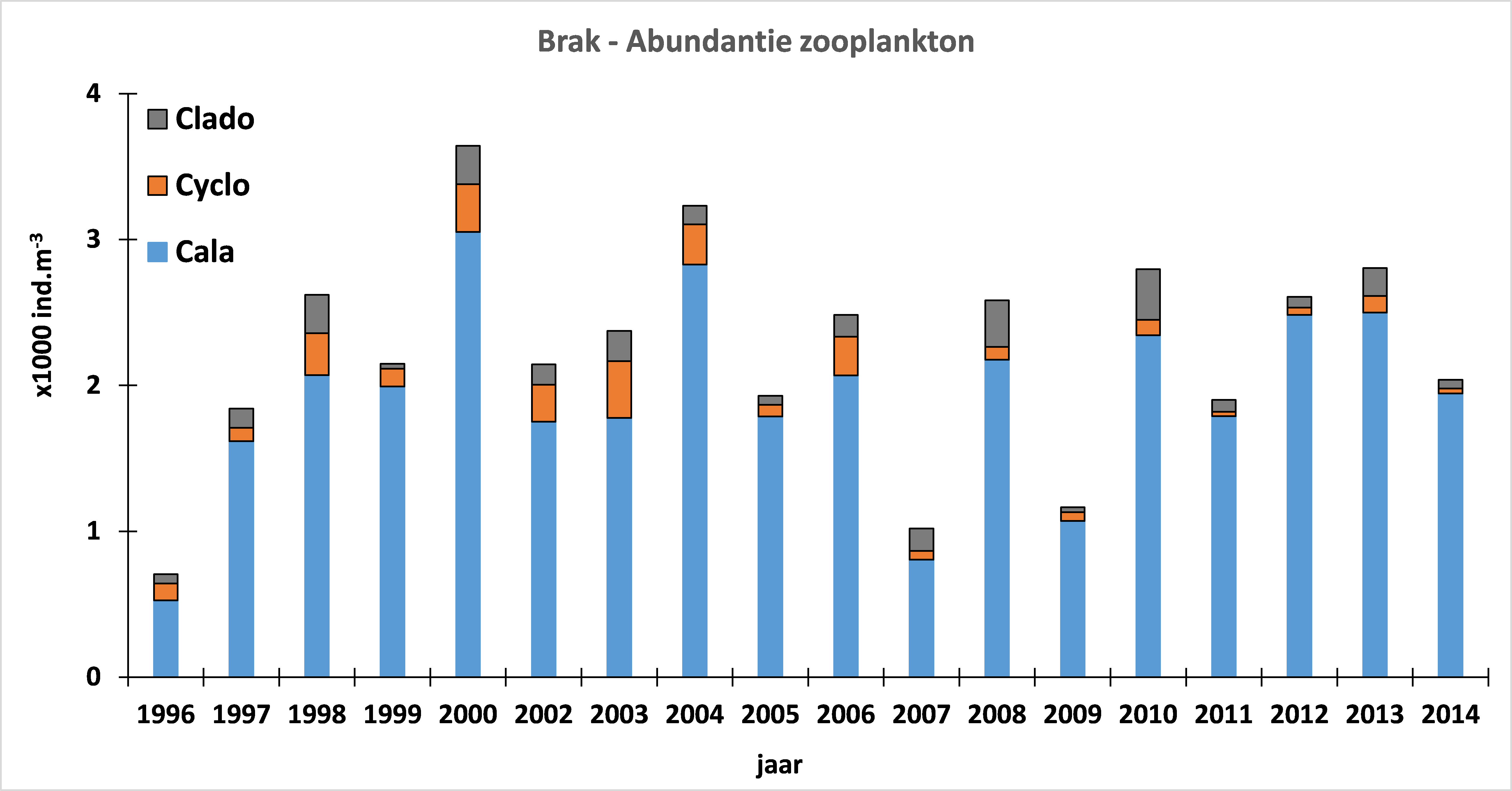
Figuur 6-45 Abundantie van mesozoöplankton organismen in het brakwater traject (Z4) tussen 1996 en 2014.
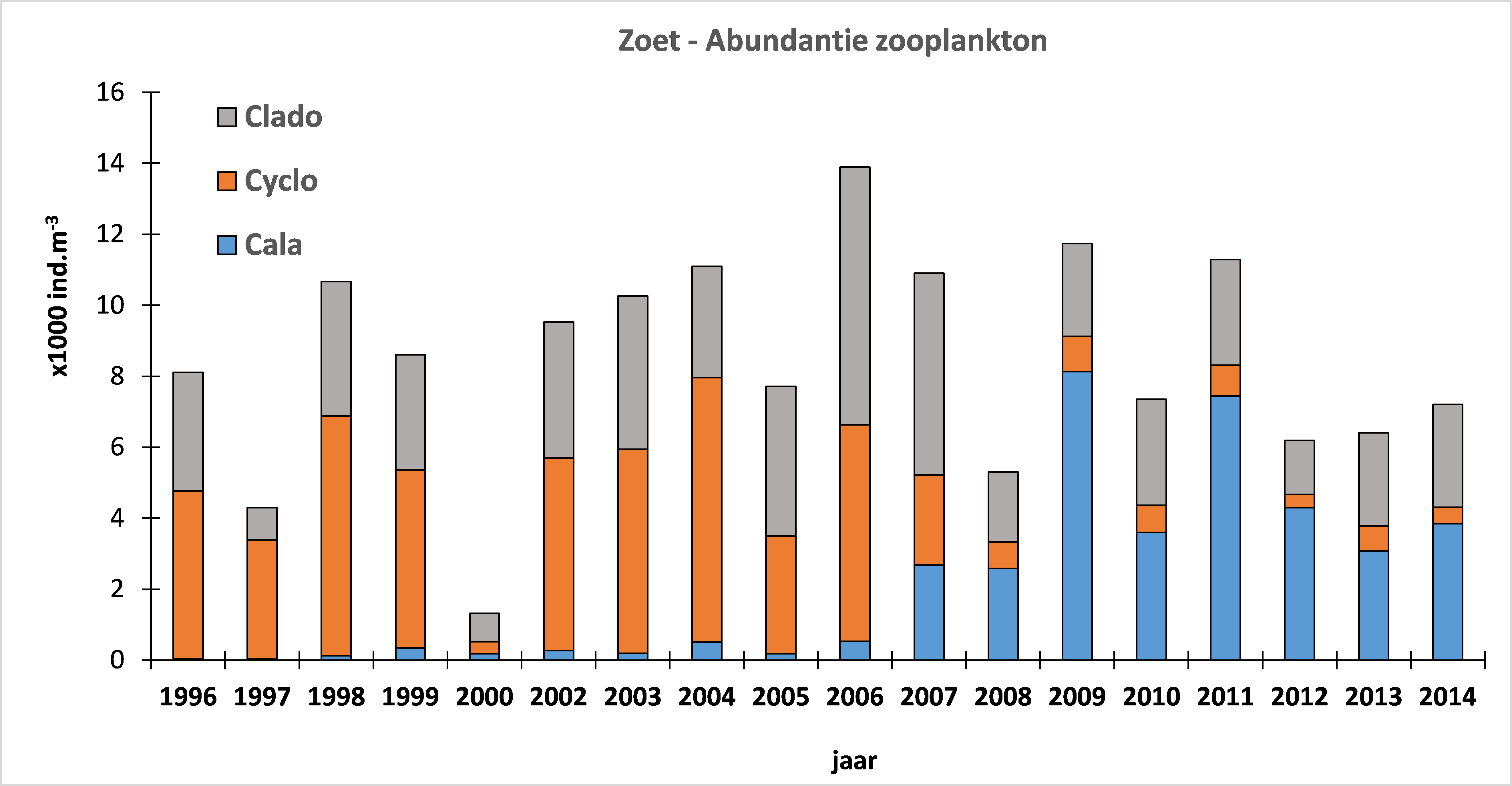
Figuur 6-46 Abundantie van mesozoöplankton organismen in het zoetwater traject (inclusief oligohalien) tussen 1996 en 2014 (Z5-Z7). In 2000 werd enkel gedurende het voorjaar bemonsterd.
In het brakwater traject varieert de jaarlijks gemiddelde totale mesozooplankton abundantie tussen 700 en 3600 ind. m-3. Calanoide copepoden domineren sterk, cyclopoide copepoden en cladoceren zijn weinig abundant. Er is geen duidelijk tendens in abundantie van deze groepen in de loop van de tijd. De abundantie (4300-7400 ind. m-3) en diversiteit aan zoöplankton is veel hoger in de zoete Zeeschelde dan in het brakwater traject. Eurytemora affinis, een copepode soort die voorheen alleen in het brakwater voorkwam, heeft er zich sterk ontwikkeld sinds 2007 en blijft domineren tot in 2014. De oorspronkelijk abundante cyclopoide copepoden zijn sterk in abundantie afgenomen. Cladoceren zijn ongewijzigd gebleven in abundantie.
Uit de RDA en GLM analyses (Tabel 6-7; Tabel 6-8) blijkt dat de toegenomen zuurstofconcentratie, gecombineerd met een dalende NH4 concentratie de belangrijkste factoren zijn die de sterke ontwikkeling van E. affinis in de stroomopwaartse Zeeschelde kunnen verklaren. Daarbij manifesteren zich duidelijk drempel effecten: de soort heeft een minimun zuurstofconcentratie van 5 mg L-1 nodig (Figuur 6-47a), en verdraagt geen NH4-N concentraties hoger dan 1 mg L-1 (Figuur 6-47b). Op de figuur staat de grenslijn op ca 5 mg/l zuurstof en 1 mg/l NH4-N?
Tabel 6-7 Resultaten van de RDA analyse voor het testen van de onafhankelijke bijdrage van verschillende omgevingsfactoren in het verklaren van de spatio-temporele abundantie van de zoöplankton gemeenschap in zone 6-7 (Dendermonde, Uitbergen, Melle). De factoren zijn gerangchikt volgens eigenwaarde (λ), *, **, *** : p< 0.05, p< 0.01, p < 0.001 significantie niveau, volgens Monte Carlo permutatie test.
| RDA conditional effects | |||
| Factors | λ | F-value | p-value |
| T | 0.1 | 43.72 | 0.001*** |
| O2 | 0.09 | 35.42 | 0.001 *** |
| BOD5 | 0.03 | 11.16 | 0.001*** |
| CL | 0.02 | 8.97 | 0.001*** |
| NH4-N | 0.01 | 6.4 | 0.001*** |
| Q | 0.02 | 5.86 | 0.001*** |
| Chla | 0.01 | 5.41 | 0.003** |
| SPM | 0 | 3.41 | 0.010** |
| NO3-N | 0.01 | 2.53 | 0.040* |
| NO2-N | 0 | 3.02 | 0.014* |
| Dsi | 0.01 | 2.6 | 0.031* |
| SO4 | 0 | 2.43 | 0.042* |
| PO4-P | 0.01 | 2.01 | 0.079 |
| DOC | 0.01 | 1.53 | 0.153 |
| pH | 0 | 1.63 | 0.155 |
| Ptot | 0 | 1.31 | 0.235 |
| Ntot | 0 | 1.29 | 0.262 |
| KJEHLN | 0 | 1.15 | 0.312 |
| Conductivity (Cond) | 0 | 1.13 | 0.361 |
| Sum of all λ | 0.32 | ||
Tabel 6-8 Resultaten van de hierarchical partitioning en GLM analyse voor E. affinis, cyclopoiden en cladoceren gedurende de Winter, lente en zomer. De relative onafhankeleijke bijdrage van ieder omgevingsfactor als predictor van de distributie van elke zooplankton groep is uitgedrukt als percent van de totale onafhankelijke bijdrage. Significantie is aangeduid als *, **, *** voor p< 0.05, 0.01 en 0.001 (ANOVA)
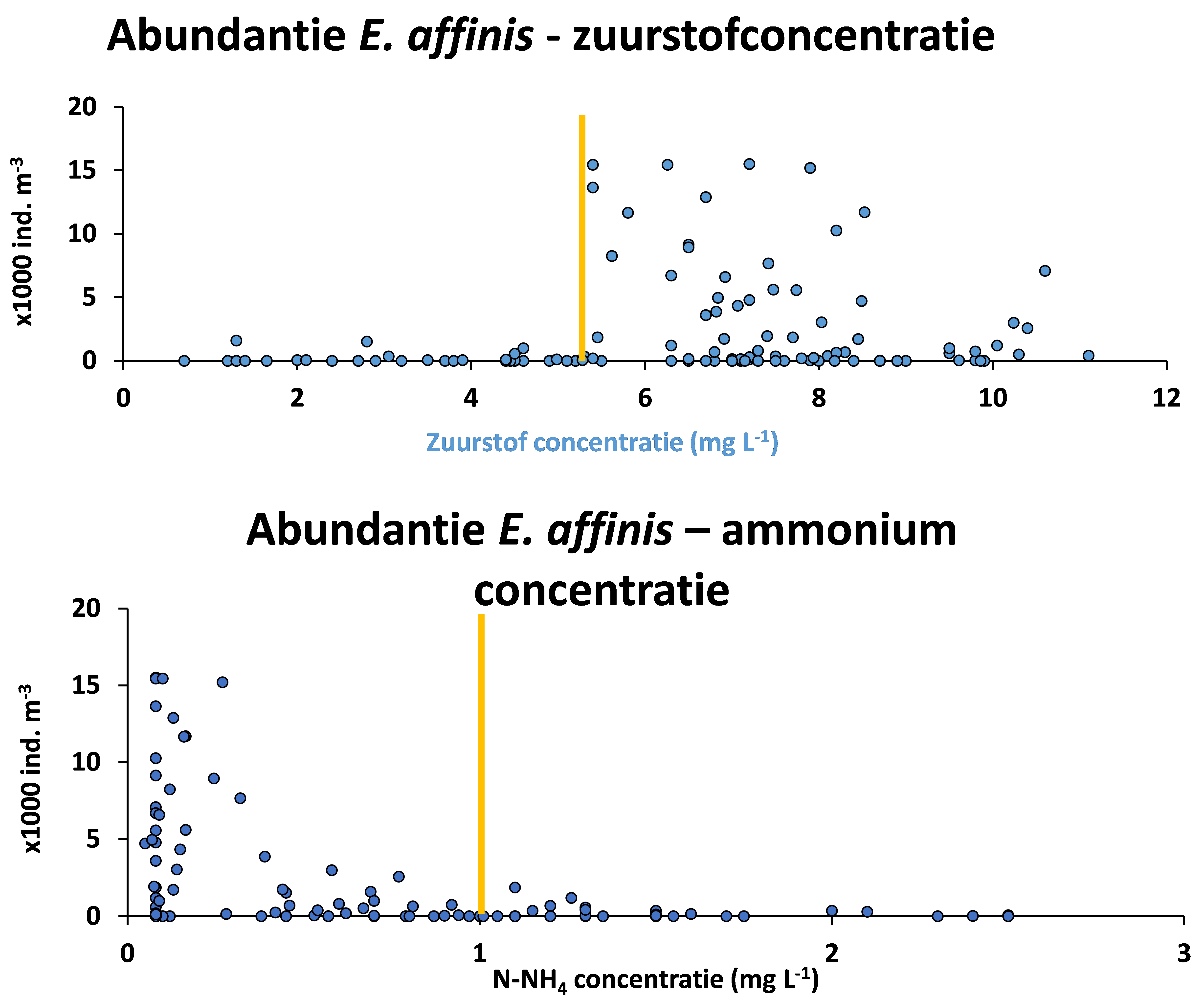
Figuur 6-47 E. affinis abundantie in de stroomopwaartse zone (Dendermonde – Melle) van de Schelde gedurende 2002-2012 in functie van de zuurstofconcentratie (a) en van de NH4-N concentratie (b).
Opmerkelijk is dat E. affinis zich in de Zeeschelde maximaal ontwikkelt bij hogere temperaturen (gemiddeld 18.58°C bij Melle) dan in de brakwaterzone, waar de te temperatuur gemiddeld 9.56°C bedraagt (station 4, Grens (Z4)).
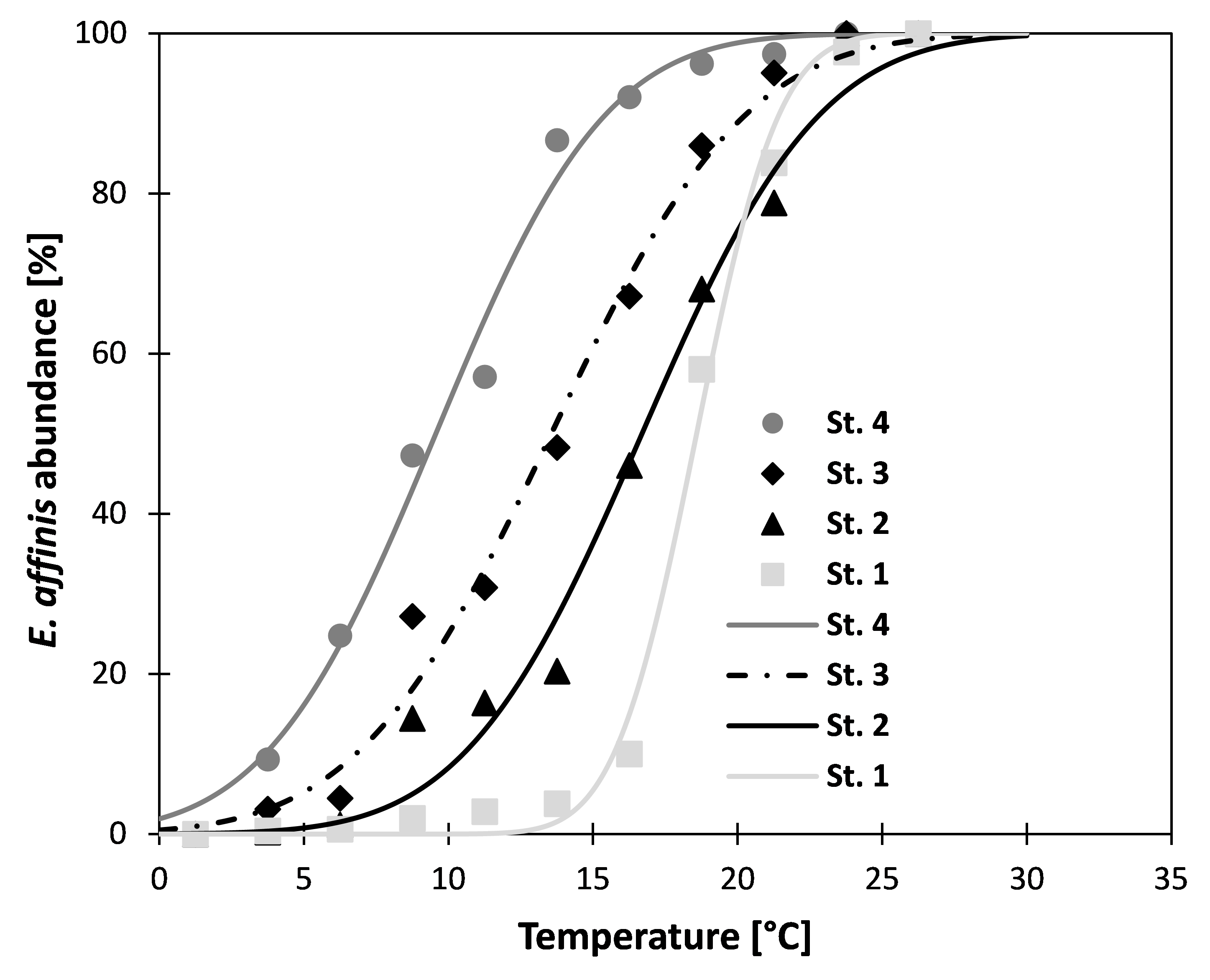
Figuur 6-48 Cumulatieve distributie van gemiddelde abundantie van E. affinis in functie van temperatuurklassen. Grens (St. 4), Dendermonde (St. 3), Uitbergen (St. 2) and Melle (St. 1). Geobserveerde data (symbolen) werden gefit met een normale cumunatieve probabiliteits- functie (lijnen).
Tabel 6-9 Parameter waarden voor de normale cumulatieve probabiliteits-functie voor gemiddelde E. affinis abundantie statistische gegevens over deze fit.
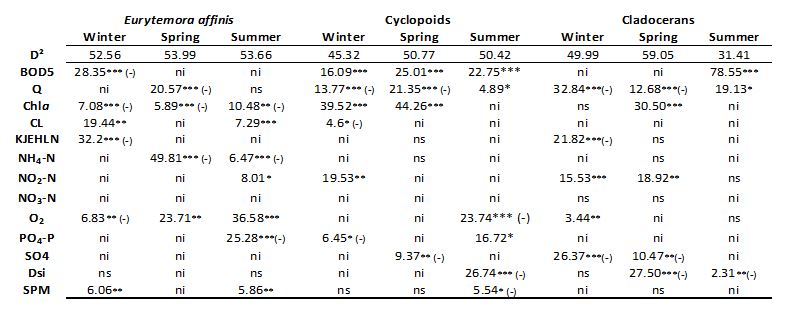
Ook binnen de zone Dendermonde – Melle schuift de gemiddelde temperatuur overeenkomend met maximale E. affinis abundantie op van 13,56 °C te Dendermonde naar 18,58 °C te Melle. Deze verschuiving kan worden verklaard doordat de waterkwaliteit in Uitbergen en Melle later op het jaar voldoet, tenminste voor NH4-N aan de normen waarbij E. affinis zich kan ontwikkelen dan in Dendermonde of in de brakwaterzone.
Deze waarneming is bijzonder interessant omdat ze een aantoont dat E. affinis, die in alle gematigde estuaria voorkomt als een typische lentesoort, een grotere plasticiteit in temperatuur tolerantie heeft dan tot hiertoe werd vermoed.
Anderzijds is geen éénduidige verklaring te vinden voor de sterke daling in abundantie van cyclopoide copepoden. Duidelijk is dat deze organismen het beter doen bij slechtere waterkwaliteit dan momenteel in de Zeeschelde voorhanden is. De huidige hypothese is dan ook dat het afnemen van de BOD5, die een afname in concentratie aan organisch materiaal vertegenwoordigt, negatief is voor deze organismen, die zich waarschijnlijk hoofdzakelijk via de microbiële loop voeden. Cladoceren blijven vrij constant in abundantie en blijken dus vrij onafhankelijk van de waterkwaliteit.
6.6.2.Grazing experimenten
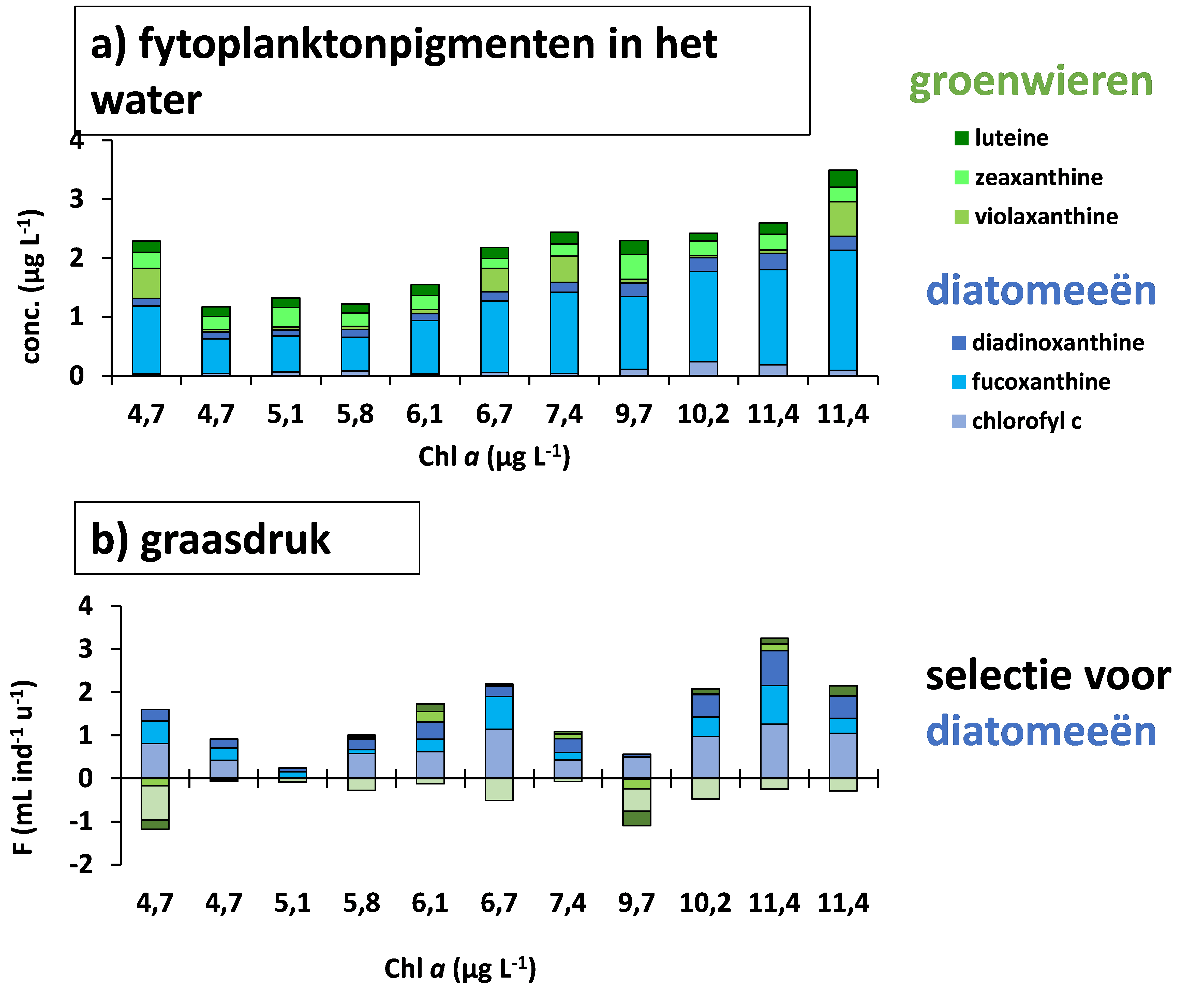
Figuur 6-49 grazing experimenten: a) fytoplankton in het water; b) graasdruk
Figuur 6-49 toont de concentratie van de belangrijkste gedetecteerde fytoplankton-merker pigmenten in de 11 grazing-experimenten uitgevoerd in April 2013 en April 2014, gerangschikt volgens stijgende chlorofyl a concentratie. Hieruit blijkt dat diatomeeën steeds domineren, zoals ook aangetoond door de microscopische tellingen. De graasdruk uitgeoefend door E. affinis adulten is systematisch hoger op diatomeeën dan op groenwieren. We benadrukken dat de berekening van de graasdruk volgens Frost (1972) een maat is voor selectie van voeding, wat betekent dat diatomeeën disproportioneel veel worden gegeten ten opzichte van hun relatieve abundantie in het milieu. De berekende graasdruk uitgeoefend door de totale E. affinis populatie is laag: tussen 4% dag-1 op de totale fytoplankton biomassa, en 6 % op de diatomeeën. Hierbij moet worden opgemerkt dat gut – content analyse uitgevoerd op E. affinis ook een substantiële ingestie van cryptophyten laat zien, die we niet in de pigmentanalyse van de incubatie experimenten terugvinden. Meer gedetailleerde analyse van de chromatogrammen uit de incubatie experimenten is aan de gang om deze verschillende resultaten te verklaren.
Referenties
De Pauw, N., 1973. On the distribution of Eurytemora affinis (Poppe, 1880) (Copepoda) in the Westerschelde estuary. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 18, 1462e1472.
Frost, B.W., 1972. Effects of size and concentration of food particles on the feeding behaviour of the marine planktonic copepod Calanus pacificus.--Limnol.Oceanogr.,17: 805-815.
Mackas D. and R. Bohrer, 1976. Fluorescence analysis of zooplankton gut contents and an investigation of diel feeding pattern. J. Exp. Biol. Ecol., 25: 77-85
Mialet, B., J. Gouzou, F. Azémar, T. Maris, C. Sossou, N. Toumi, S. Van Damme, P. Meire, and M. Tackx. 2011. Response of zooplankton to improving water quality in the Scheldt estuary (Belgium). Estuarine, Coastal and Shelf Science 93:47-57.
Tackx M., S. Chambord, F AzémarC. Sossou, M. Le Coz & R. Fernandez, 2015.
Onderzoek naar de gevolgen van het Sigmaplan, baggeractiviteiten en havenuitbreiding in de Zeeschelde op het milieu. OMES verslag 2013-’14. 42 pp.
Hoofdstuk 7.Conclusie
De voorbije 20 jaar heeft de Zeeschelde een enorme evolutie doorgemaakt. Eind vorige eeuw had de Zeeschelde de kwalijke reputatie van quasi dode rivier. Lozingen van onbehandeld huishoudelijk en industrieel afvalwater en grote run-off uit landbouwgebieden waren de grote boosdoeners. Het water bevatte hoge vrachten aan nutriënten (N en P). De afbraak van organische koolstofverbindingen en de nitrificatie van ammonium onttrokken veel zuurstof aan het water. De gevolgen van de toenemende vervuiling leidden tot de vernietiging van het aquatische ecosysteem in de zoete Zeeschelde. In de zomer verdween quasi alle zuurstof naar de mineralisatie van de organische koolstofvracht (BOD) en de nitrificatie van NH4+. De steeds toenemende vuilvracht oversteeg echter het zelf zuiverende vermogen van de zoete Schelde: de zoete zone krijgt niet meer alle BOD en NH4+ verwerkt, zodat steeds grotere vrachten naar afwaartse zones worden doorgegeven, zodat ook in de Beneden-Zeeschelde het zuurstofgehalte ’s zomers onder 2 mg/l zakte.
Vanaf dan worden acties ondernomen om de instroom van vervuiling naar het estuarium te beperken. Industrie en landbouw krijgen stelselmatig strengere normen opgelegd, huishoudens worden meer en meer aangesloten op waterzuiveringsinstallaties. In de jaren ’70 was waterzuivering quasi onbestaande in het Scheldebekken. In 1990 bedroeg de zuiveringsgraad slechts 30%. Vanaf dan werd waterzuivering een gewestelijke bevoegdheid in Vlaanderen en werd de naamloze vennootschap Aquafin opgericht. Er startte een inhaalbeweging: in 2000 bedroeg de zuiveringsgraad 52%, in 2008 73%. Sinds 2007 wordt ook het leeuwendeel van het afvalwater uit het Brusselse Gewest dat voor kort nog ongezuiverd via Zenne, Dijle en Rupel in de Schelde kwam, gezuiverd. Ook in Wallonië en Frankrijk stijgt de capaciteit aan rioolwaterzuiveringsinstallaties.
De gevolgen van deze inspanningen zijn duidelijk merkbaar. De vuilvracht vanuit het bekken neemt af, en het zuurstofgehalte in de Zeeschelde stijgt. Opmerkelijk is wel dat deze stijging niet altijd mooi parallel verloopt met de toename aan waterzuiveringscapaciteit. Door de vernietiging van het aquatische ecosysteem zat de zoete Zeeschelde als het ware in een zuurstofloze wurggreep. Ondanks een gestage verbetering in het bekken, bleef de Zeeschelde ’s zomers een bijna zuurstofloos systeem. Tot ca 2003, wanneer een keerpunt wordt bereikt. Een verbeterde waterkwaliteit maakt sterke algenbloei terug mogelijk. Voordien was immers zelfs algenbloei geremd door de slechte waterkwaliteit. Maar eens de algenbloei terug hoge concentraties bereikt, schiet zuurstof weer de hoogte in. Het ecosysteem kan herademen en zorgt zelf voor een verdere verbetering van de waterkwaliteit. Op amper enkele jaren tijd slaat het systeem om van een quasi zuurstofloos naar een zuurstofrijk systeem. Dankzij deze verbeterde waterkwaliteit kan ook het andere leven herademen in de Zeeschelde. De laatste jaren wordt op de meeste plaatsen in het estuarium de zuurstofnorm uit de Evaluatiemethodiek gehaald. Enkel in de zijriveren Durme en Rupel, en in de oligohaliene zone waar ze in uitmonden, was er anno 2015 een beperkt tekort.
De Schelde is nu wel zuurstofrijk, ze is verre van een volledig gezond systeem. Ze evolueerde van een oververvuild naar een klassiek vervuild of eutroof systeem. Er is nog steeds een te grote biochemische zuurstofvraag, in het bijzonder voor de nitrificatie van een overmaat aan ammonium. Ook fosfaat en nitriet voldoen nog niet aan de criteria uit de Evaluatiemethodiek. In het klassieke eutrofiëringsverhaal leiden verhoogde concentraties aan stikstof en fosfor tot buitensporige algenbloei. Tot dergelijke bloei, met de klassieke neveneffecten van nachtelijke zuurstoftekorten, het ontstaan van schuimvorming of toxische stoffen, leidt de overmaat aan N en P in de Schelde echter niet. Het is lichtgebrek dat wellicht de algenbloei onderdrukt. Estuaria zoals de Schelde worden gekenmerkt door een hoge troebelheid waardoor zonlicht niet diep kan doordringen en de eufotische diepte (belichte zone) beperkt blijft. Maar naast eufotische diepte (Zeu), speelt ook de mengdiepte (Zm) een rol: hoe dieper, hoe langer een alg in het donker zal zijn en niet aan fotosynthese kan doen. De verhouding fotische diepte/mengdiepte is een goede maat voor de lichtlimitatie. Wanneer lichtlimitatie optreedt, kunnen algen het nutriëntaanbod niet ten volle benutten waardoor een grote nutriëntvracht kan doorstromen naar afwaartse gebieden en de kustzee.
Hoewel de meest opwaartse delen van de Zeeschelde de grootste troebelheid kennen (= beperkte fotische diepte), is door de geringe mengdiepte het lichtklimaat hier toch redelijk gunstig. Deze zone kent bijgevolg de hoogste primaire productie. De maximum fotosynthese snelheid (PBmax) van het fytoplankton lijkt vooral af te hangen van de temperatuur terwijl de primaire productie per eenheid biomassa (PB-ratio) een goede relatie vertoonde met de Zeu/Zm-ratio. Dit wijst er sterk op dat de primaire productie van het fytoplankton vooral wordt bepaald door de lichtbeschikbaarheid van het water. Dus alles wat dit lichtklimaat beïnvloedt zal een weerslag hebben op de primaire productie en daarmee op de potentiele draagkracht van het gehele Schelde ecosysteem.
In een gezond ecosysteem wordt algenbloei niet louter bepaald door lichtbeschikbaarheid, maar zorgt begrazing door zoöplankton voor een regulering van de algenbloei. De aanhoudende verbetering van de waterkwaliteit in de Zeeschelde brengt met zich mee dat de zoöplankton gemeenschap in volle evolutie is en nu kan worden geobserveerd in omgevingsomstandigheden die anders (beter) zijn dan tijdens de oudste kwantitatieve zoöplankton analyses ooit in de Schelde gedaan. Tijdens de periode 1967-’69 werd het zoöplankton over de gehele Schelde bestudeerd door De Pauw (1975). Deze rapporteert dat in die periode de zuurstofconcentratie in de Zeeschelde meestal < 4 mg L-1 was, en in die periode kwam E. affinis, een belangrijke indicatorsoort in estuaria, voornamelijk in het brakwatertraject van de Schelde voor. Deze toestand is dus vergelijkbaar met de periode voor 2007. Het feit dat de waterkwaliteit in de Schelde geëvolueerd is tot het huidig niveau is dus niet alleen een beleidssucces in termen van milieukwaliteit, maar opent ook nieuwe horizonten voor het ontdekken van relaties tussen milieu en biotoop.
Gezien de dominante meso-zoöplankton organismen (E. affinis) een beperkte graasdruk op de fytoplankton gemeenschap uitoefenen, kan men stellen dat het zoöplankton compartiment waarschijnlijk geen ‘bottleneck’ vormt voor de ontwikkeling van de hogere trofische niveaus in de pelagiale Schelde voedselketen. De vraag rijst of de mesozooplankton gemeenschap eventuele blooms van algen (of cyanobacteriën) zou kunnen controleren. Waarschijnlijk dient hiervoor eerder op het microzooplankton (rotiferen, cilliaten, heterotrofe flagellaten) worden gerekend.
Om die doorstroming van biomassa en energie van algen naar zoöplankton te beoordelen, werd in de Evaluatiemethodiek de PB:B ratio voorgesteld. Deze ratio geeft de verhouding weer tussen de primaire productie (per eenheid biomassa) en de biomassa aan algen. Een hoge ratio is ideaal, dit betekent immers dat de algen zeer productief zijn maar toch beperkt zijn in biomassa. Dit kan als de algen vlot opgegeten worden en er dus een goede doorstroming is van massa/energie in de voedselketen.
De PB:B-ratio bereikt minimale waarden bij Appels en Uitbergen en de hoogste waarde bij de grens. Dit wijst erop dat nabij de grens de algen productief zijn, maar dat er geen grote biomassa wordt opgebouwd omdat de algen snel worden begraasd. De chl a concentraties zijn nabij de grens dan ook laag. Nabij Appels en Uitbergen ligt het chlorofylgehalte ruim een factor 10 hoger. De primaire productie ligt er hoger, maar de graasdruk is wellicht beperkter. Ondanks de zeer hoge chl a concentraties, is de zoöplanktondichtheid hier in het zoete slechts een factor 3 hoger dan in het brakke. De doorstroming van energie en biomassa in de voedselketen is dus minder efficiënt en scoort onvoldoende.
Het lichtklimaat werd reeds aangehaald als de sturende factor voor algenbloei en bij uitbreiding het gehele ecosysteem van de Zeeschelde. Dit lichtklimaat wordt bepaald door de troebelheid, de zwevende stof in de waterkolom. Daarom verdient de evolutie van zwevende stof bijzondere aandacht.
Op kleine schaal variëren de concentraties aan zwevende stof over de rivierbreedte, diepte (verticale) en over getijcycli. De 13-uursmetingen illustreren de getijvariatie, waar uit we algemeen kunnen stellen dat de hogere zwevende stofconcentraties voorkomen bij hogere stroomsnelheden zowel bij vloed als bij eb. Bij kentering zijn de stroomsnelheden en concentraties het laagst. Naarmate de stroomsnelheden stijgen, nemen de concentraties toe en wanneer de maximum stroomsnelheden zijn bereikt, nemen de concentraties terug af. De variaties omwille van de riviermorfologie worden voornamelijk afwaarts van Antwerpen waargenomen waar voornamelijk hogere concentraties worden vastgesteld langsheen de profielen dan gemiddeld over de gevaren dwarssectie. Al dient opgemerkt te worden dat aan station Lippenbroek de gemiddelde concentraties gemeten (tijdens eb) en over een profiel ca. 25% lager zijn dan gemeten over de dwarsraai.
De verdeling van de zwevende stof over de waterkolom vertoont een verticale gradiënt die het sterkst is in het afwaartse deel van de Zeeschelde. De verticale gradiënt neemt af in opwaartse richting doorheen de Zeeschelde; nabij de opwaartse rand is de concentratie bijna homogeen over de waterkolom. In de Beneden-Zeeschelde is de verticale gradiënt afgenomen doorheen de tijd sinds ca. 2005 (zwevende stof werd meer uniform over de waterkolom). Dit komt voornamelijk door een stijging van de concentraties in de bovenste helft van de waterkolom.
In de analyse van de oppervlaktestalen werd gesteld dat er sinds 2009 een grotere variatie is in de zomergemiddelde sedimentconcentratie in de zone tussen 100 en 130 km van de monding. De dieptestalen lijken dit te bevestigen noch te ontkrachten: er wordt wel een toename van de variatie waargenomen, maar deze is vrij zwak. Tijdens de wintermaanden ontwikkelt zich een turbiditeitsmaximum in de Beneden-Zeeschelde (ca. km 60-80). Dit maximum is sterk toegenomen sinds 2009, en dan vooral in de bovenste helft van de waterkolom. Vooral de meetlocaties Antwerpen, Liefkenshoek en Punt van Melsele vertonen een toename in de wintergemiddelde sedimentconcentratie sinds 2009, met name in de bovenste helft van de waterkolom.
De jaar- en systeemgemiddelde zwevende stofconcentraties (Boven-Zeeschelde, Beneden-Zeeschelde en Rupel) vertonen geen duidelijke trend in functie van de tijd. In de oppervlakte- en dieptestalen werden in 2004, 2009 en 2011 sterk verhoogde jaargemiddelde concentraties waargenomen.
Wel werden duidelijke relaties gevonden tussen zwevende stof en het zoetwaterdebiet: hoe lager het debiet, hoe meer zwevende stof zich accumuleert in de zoete Zeeschelde. Opvallend is dat sinds 2009, de Schelde veel gevoeliger geworden is aan lage debieten: bij lage debieten werden in de zoete Zeeschelde veel hogere zwevende stofconcentraties waargenomen dan voor 2009. Deze wijziging deed zich voor in de bovenste en de onderste helft van de waterkolom, en zowel tijdens de zomer- als tijdens de winterperiode.
Er is ook een relatie tussen de stortvolumes van fijn sediment en zwevende stof afwaarts van de stortzone (aan Antwerpen en Kruibeke) waargenomen. Aan station Liefkenshoek wordt ook een (zwakke) relatie waargenomen en is mogelijk gevolg van de baggeractiviteiten rondom Deurganckdok die vertegenwoordigd wordt in de stortvolumes. In de Boven-Zeeschelde is de relatie minder duidelijk. Indien meetjaar 2011 niet beschouwd wordt, wordt in de Boven-Zeeschelde eerder een omgekeerde relatie vastgesteld. De verhoogde concentraties in meetjaren 2004, 2009 en 2011 zijn mogelijk een gevolg van gecombineerde effecten van lage bovenafvoeren en hoge stortvolumes. De zwevende stofconcentraties zijn na 2011 terug afgenomen en dit voornamelijk in de Boven-Zeeschelde (op basis van de dieptestalen).
 OMES is a project funded by the Flemish Government
OMES is a project funded by the Flemish Government